

TLR2-mediated activation of innate responses in the upper airways confers antiviral protection of the lungs
- PMID: 33561017
- PMCID: PMC8021123
- DOI: 10.1172/jci.insight.140267
TLR2-mediated activation of innate responses in the upper airways confers antiviral protection of the lungs
Abstract
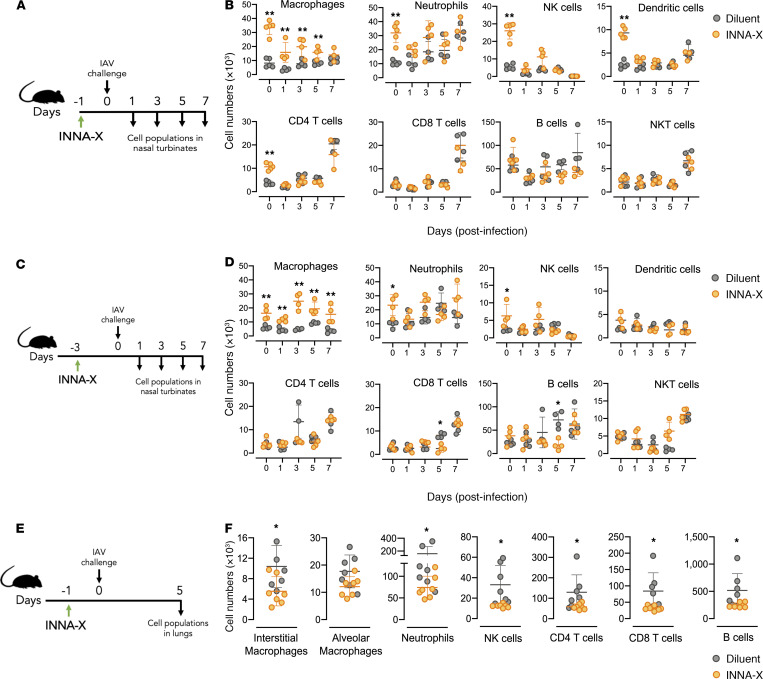
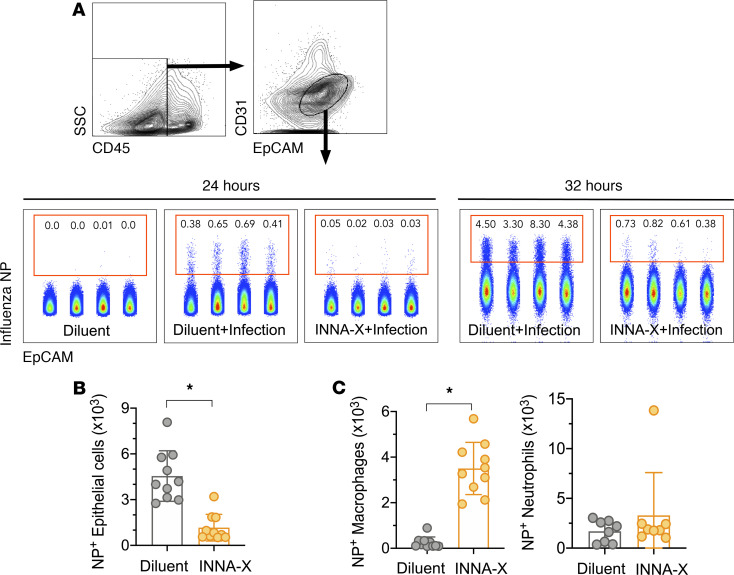
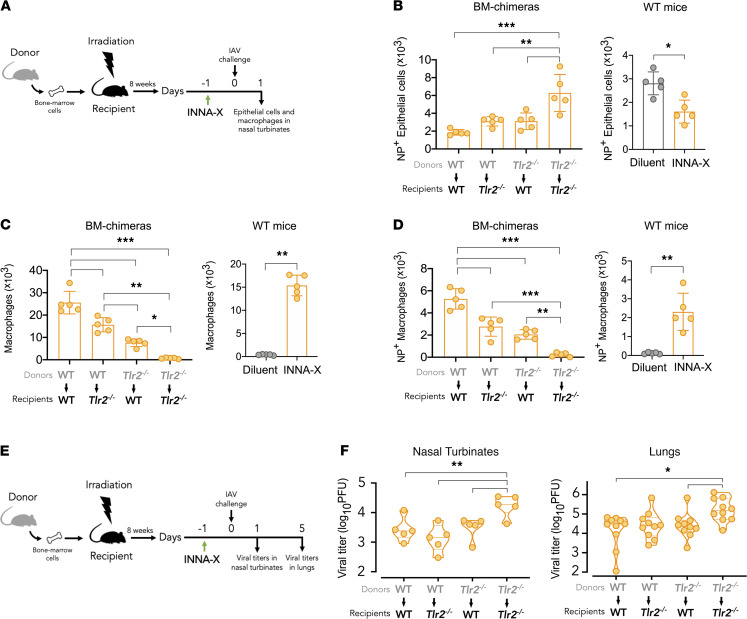
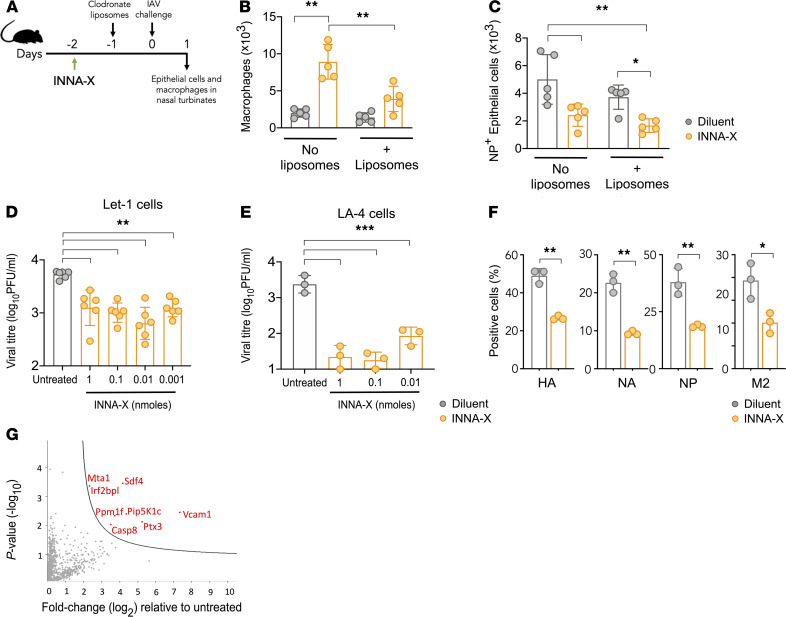
The impact of respiratory virus infections on global health is felt not just during a pandemic, but endemic seasonal infections pose an equal and ongoing risk of severe disease. Moreover, vaccines and antiviral drugs are not always effective or available for many respiratory viruses. We investigated how induction of effective and appropriate antigen-independent innate immunity in the upper airways can prevent the spread of respiratory virus infection to the vulnerable lower airways. Activation of TLR2, when restricted to the nasal turbinates, resulted in prompt induction of innate immune-driven antiviral responses through action of cytokines, chemokines, and cellular activity in the upper but not the lower airways. We have defined how nasal epithelial cells and recruitment of macrophages work in concert and play pivotal roles to limit progression of influenza virus to the lungs and sustain protection for up to 7 days. These results reveal underlying mechanisms of how control of viral infection in the upper airways can occur and support the implementation of strategies that can activate TLR2 in nasal passages to provide rapid protection, especially for at-risk populations, against severe respiratory infection when vaccines and antiviral drugs are not always effective or available.
Keywords: Infectious disease; Influenza; Innate immunity; Therapeutics.
Conflict of interest statement
Figures
Similar articles
-
Inactivated Influenza Vaccine That Provides Rapid, Innate-Immune-System-Mediated Protection and Subsequent Long-Term Adaptive Immunity.mBio. 2015 Oct 27;6(6):e01024-15. doi: 10.1128/mBio.01024-15. mBio. 2015. PMID: 26507227 Free PMC article.
-
Intranasal administration of the TLR2 agonist Pam2Cys provides rapid protection against influenza in mice.Mol Pharm. 2012 Sep 4;9(9):2710-8. doi: 10.1021/mp300257x. Epub 2012 Aug 3. Mol Pharm. 2012. PMID: 22823162
-
IFN-λ prevents influenza virus spread from the upper airways to the lungs and limits virus transmission.Elife. 2018 Apr 13;7:e33354. doi: 10.7554/eLife.33354. Elife. 2018. PMID: 29651984 Free PMC article.
-
Innate Immune Responses to Influenza Virus Infections in the Upper Respiratory Tract.Viruses. 2021 Oct 17;13(10):2090. doi: 10.3390/v13102090. Viruses. 2021. PMID: 34696520 Free PMC article. Review.
-
Innate immunity to influenza in chronic airways diseases.Respirology. 2012 Nov;17(8):1166-75. doi: 10.1111/j.1440-1843.2012.02200.x. Respirology. 2012. PMID: 22616906 Review.
Cited by
-
Transcriptional Analysis of lncRNA and Target Genes Induced by Influenza A Virus Infection in MDCK Cells.Vaccines (Basel). 2023 Oct 14;11(10):1593. doi: 10.3390/vaccines11101593. Vaccines (Basel). 2023. PMID: 37896995 Free PMC article.
-
Influence of Donor-Specific Characteristics on Cytokine Responses in H3N2 Influenza A Virus Infection: New Insights from an Ex Vivo Model.Int J Mol Sci. 2024 Oct 11;25(20):10941. doi: 10.3390/ijms252010941. Int J Mol Sci. 2024. PMID: 39456722 Free PMC article.
-
Recent Insights into the Molecular Mechanisms of the Toll-like Receptor Response to Influenza Virus Infection.Int J Mol Sci. 2024 May 29;25(11):5909. doi: 10.3390/ijms25115909. Int J Mol Sci. 2024. PMID: 38892096 Free PMC article. Review.
-
Advances in the Prophylaxis of Respiratory Infections by the Nasal and the Oromucosal Route: Relevance to the Fight with the SARS-CoV-2 Pandemic.Pharmaceutics. 2022 Feb 27;14(3):530. doi: 10.3390/pharmaceutics14030530. Pharmaceutics. 2022. PMID: 35335905 Free PMC article. Review.
-
Understanding Rhinovirus Circulation and Impact on Illness.Viruses. 2022 Jan 13;14(1):141. doi: 10.3390/v14010141. Viruses. 2022. PMID: 35062345 Free PMC article. Review.
References
-
- Centers for Disease Control and Prevention. Past Seasons Vaccine Effectiveness Estimates. https://www.cdc.gov/flu/vaccines-work/past-seasons-estimates.html Updated January 29, 2020. Accessed February 5, 2021.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
