

Fuzzy Association of an Intrinsically Disordered Protein with Acidic Membranes
- PMID: 33554215
- PMCID: PMC7851954
- DOI: 10.1021/jacsau.0c00039
Fuzzy Association of an Intrinsically Disordered Protein with Acidic Membranes
Abstract
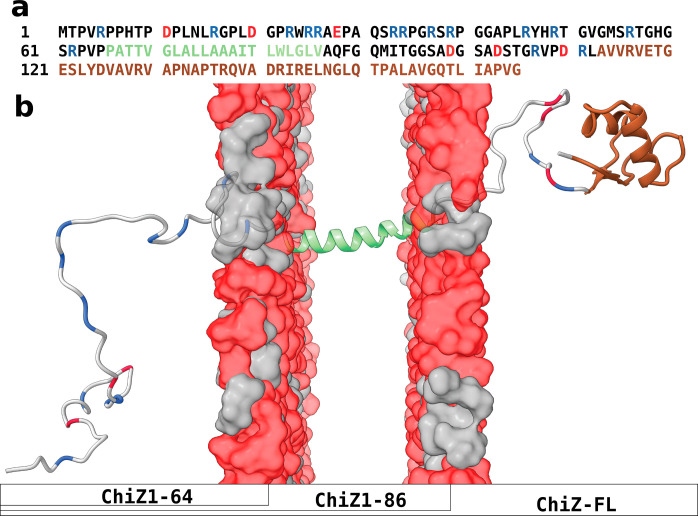
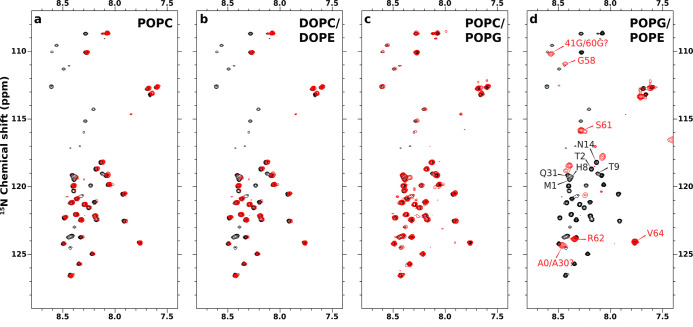
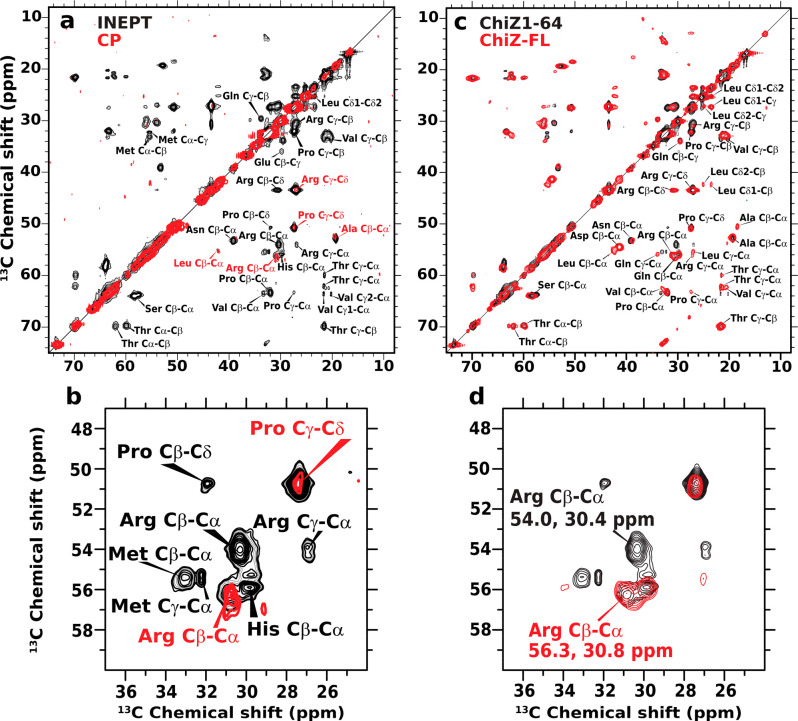
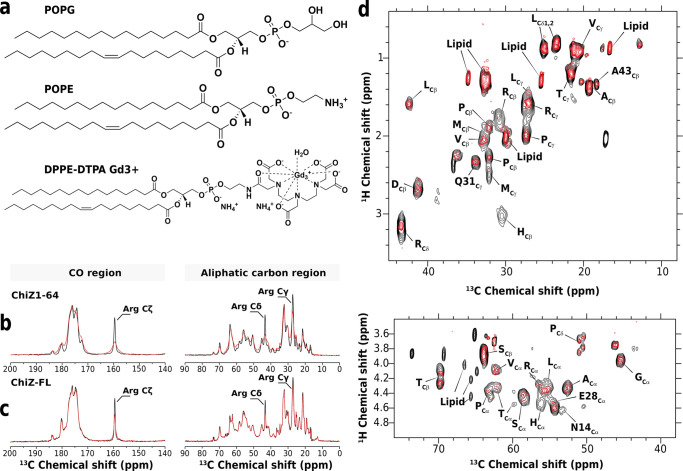
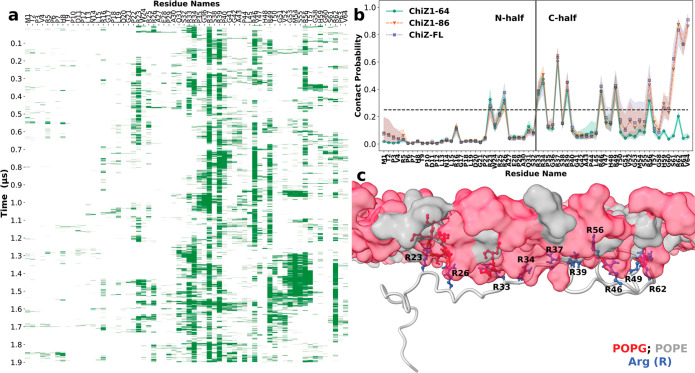
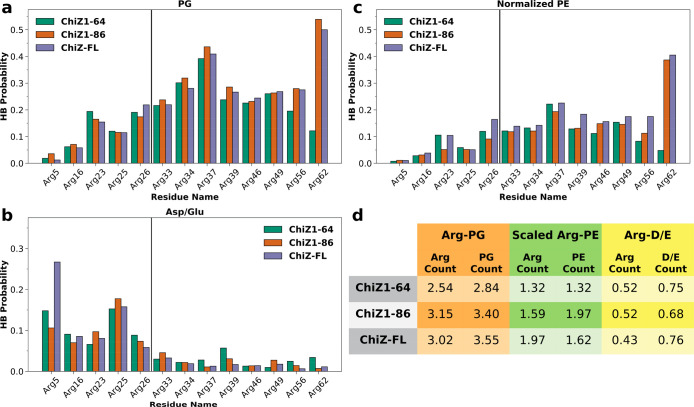
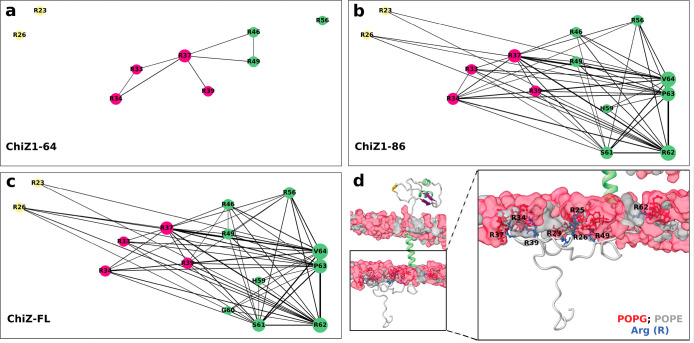
Many physiological and pathophysiological processes, including Mycobacterium tuberculosis (Mtb) cell division, may involve fuzzy membrane association by proteins via intrinsically disordered regions. The fuzziness is extreme when the conformation and pose of the bound protein and the composition of the proximal lipids are all highly dynamic. Here, we tackled the challenge in characterizing the extreme fuzzy membrane association of the disordered, cytoplasmic N-terminal region (NT) of ChiZ, an Mtb divisome protein, by combining solution and solid-state NMR spectroscopy and molecular dynamics simulations. While membrane-associated NT does not gain any secondary structure, its interactions with lipids are not random, but formed largely by Arg residues predominantly in the second, conserved half of the NT sequence. As NT frolics on the membrane, lipids quickly redistribute, with acidic lipids, relative to zwitterionic lipids, preferentially taking up Arg-proximal positions. The asymmetric engagement of NT arises partly from competition between acidic lipids and acidic residues, all in the first half of NT, for Arg interactions. This asymmetry is accentuated by membrane insertion of the downstream transmembrane helix. This type of semispecific molecular recognition may be a general mechanism by which disordered proteins target membranes.
© 2020 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
Structure of CrgA, a cell division structural and regulatory protein from Mycobacterium tuberculosis, in lipid bilayers.Proc Natl Acad Sci U S A. 2015 Jan 13;112(2):E119-26. doi: 10.1073/pnas.1415908112. Epub 2014 Dec 29. Proc Natl Acad Sci U S A. 2015. PMID: 25548160 Free PMC article.
-
Fuzzy regions in an intrinsically disordered protein impair protein-protein interactions.FEBS J. 2016 Feb;283(4):576-94. doi: 10.1111/febs.13631. Epub 2016 Jan 12. FEBS J. 2016. PMID: 26684000
-
The intrinsically disordered N-terminal domain of galectin-3 dynamically mediates multisite self-association of the protein through fuzzy interactions.J Biol Chem. 2017 Oct 27;292(43):17845-17856. doi: 10.1074/jbc.M117.802793. Epub 2017 Sep 11. J Biol Chem. 2017. PMID: 28893908 Free PMC article.
-
Extreme Fuzziness: Direct Interactions between Two IDPs.Biomolecules. 2019 Feb 26;9(3):81. doi: 10.3390/biom9030081. Biomolecules. 2019. PMID: 30813629 Free PMC article. Review.
-
Modulation of Measles Virus NTAIL Interactions through Fuzziness and Sequence Features of Disordered Binding Sites.Biomolecules. 2018 Dec 27;9(1):8. doi: 10.3390/biom9010008. Biomolecules. 2018. PMID: 30591682 Free PMC article. Review.
Cited by
-
Partial mimicry of the microtubule binding of tau by its membrane binding.Protein Sci. 2023 Mar;32(3):e4581. doi: 10.1002/pro.4581. Protein Sci. 2023. PMID: 36710643 Free PMC article.
-
NMR illuminates intrinsic disorder.Curr Opin Struct Biol. 2021 Oct;70:44-52. doi: 10.1016/j.sbi.2021.03.015. Epub 2021 May 2. Curr Opin Struct Biol. 2021. PMID: 33951592 Free PMC article. Review.
-
Control of G protein-coupled receptor function via membrane-interacting intrinsically disordered C-terminal domains.Proc Natl Acad Sci U S A. 2024 Jul 16;121(29):e2407744121. doi: 10.1073/pnas.2407744121. Epub 2024 Jul 10. Proc Natl Acad Sci U S A. 2024. PMID: 38985766 Free PMC article.
-
Illuminating Disorder Induced by Glu in a Stable Arg-Anchored Transmembrane Helix.ACS Omega. 2021 Jul 26;6(31):20611-20618. doi: 10.1021/acsomega.1c02800. eCollection 2021 Aug 10. ACS Omega. 2021. PMID: 34396006 Free PMC article.
-
Elucidating the Membrane Binding Process of a Disordered Protein: Dynamic Interplay of Anionic Lipids and the Polybasic Region.ACS Phys Chem Au. 2024 Jan 18;4(2):167-179. doi: 10.1021/acsphyschemau.3c00051. eCollection 2024 Mar 27. ACS Phys Chem Au. 2024. PMID: 38560754 Free PMC article.
References
-
- Borgia A.; Borgia M. B.; Bugge K.; Kissling V. M.; Heidarsson P. O.; Fernandes C. B.; Sottini A.; Soranno A.; Buholzer K. J.; Nettels D.; Kragelund B. B.; Best R. B.; Schuler B. Extreme disorder in an ultrahigh-affinity protein complex. Nature 2018, 555 (7694), 61–66. 10.1038/nature25762. - DOI - PMC - PubMed
-
- Neira J. L.; Palomino-Schätzlein M.; Ricci C.; Ortore M. G.; Rizzuti B.; Iovanna J. L. Dynamics of the intrinsically disordered protein NUPR1 in isolation and in its fuzzy complexes with DNA and prothymosin α. Biochim. Biophys. Acta, Proteins Proteomics 2019, 1867 (11), 140252.10.1016/j.bbapap.2019.07.005. - DOI - PubMed
-
- Meyer N. H.; Dellago H.; Tam-Amersdorfer C.; Merle D. A.; Parlato R.; Gesslbauer B.; Almer J.; Gschwandtner M.; Leon A.; Franzmann T. M.; Grillari J.; Kungl A. J.; Zangger K.; Falsone S. F. Structural fuzziness of the RNA-organizing protein SERF determines a toxic gain-of-interaction. J. Mol. Biol. 2020, 432 (4), 930–951. 10.1016/j.jmb.2019.11.014. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials