

Safety, Immunogenicity, and Protective Efficacy of a Chimeric A/B Live Attenuated Influenza Vaccine in a Mouse Model
- PMID: 33513862
- PMCID: PMC7910998
- DOI: 10.3390/microorganisms9020259
Safety, Immunogenicity, and Protective Efficacy of a Chimeric A/B Live Attenuated Influenza Vaccine in a Mouse Model
Abstract
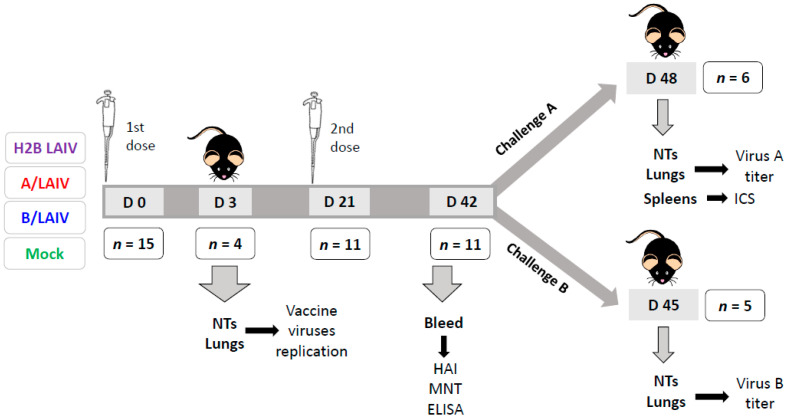
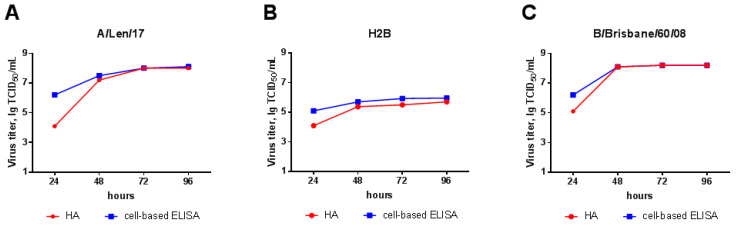
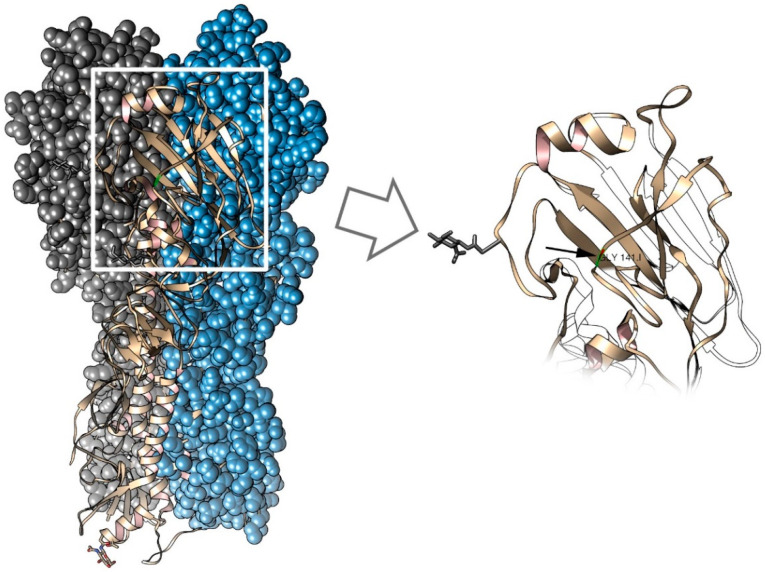
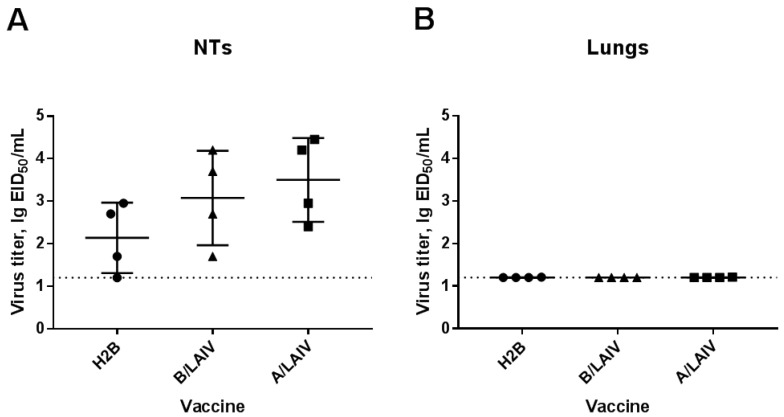
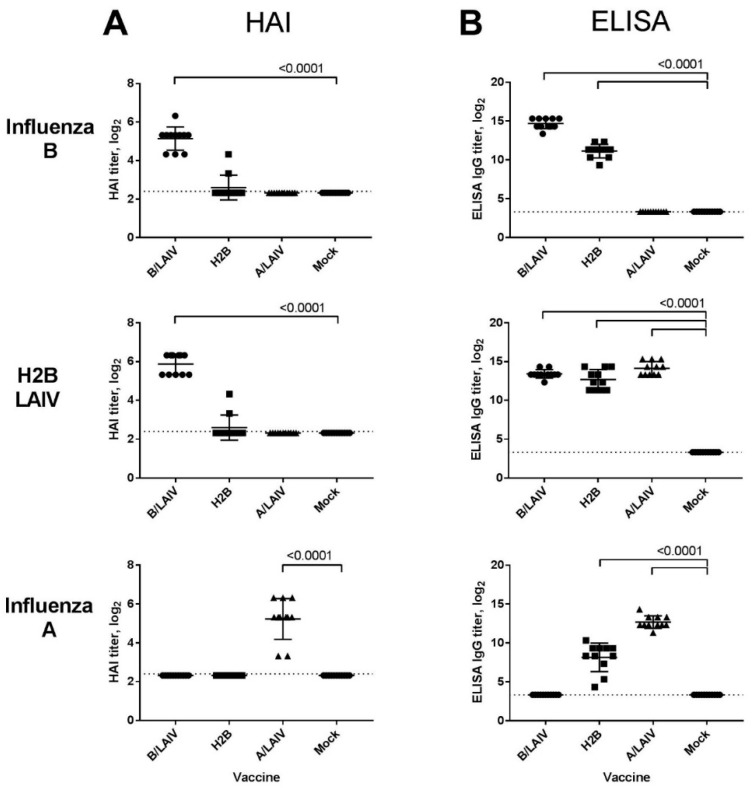
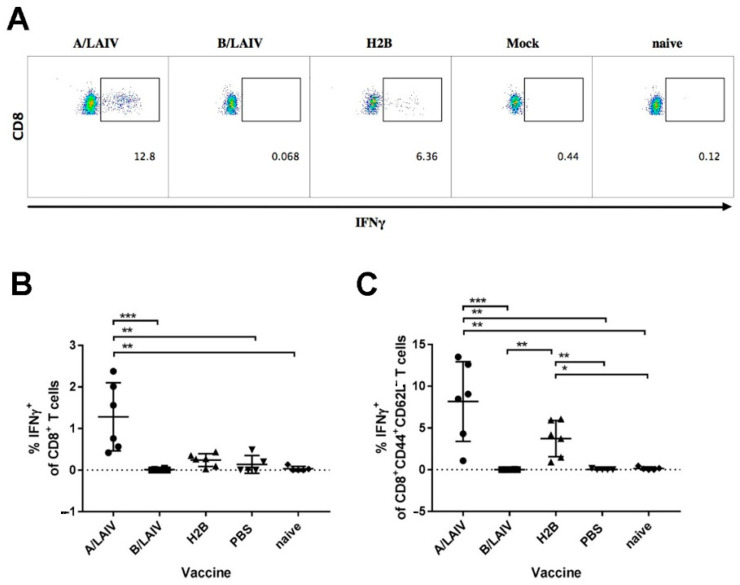
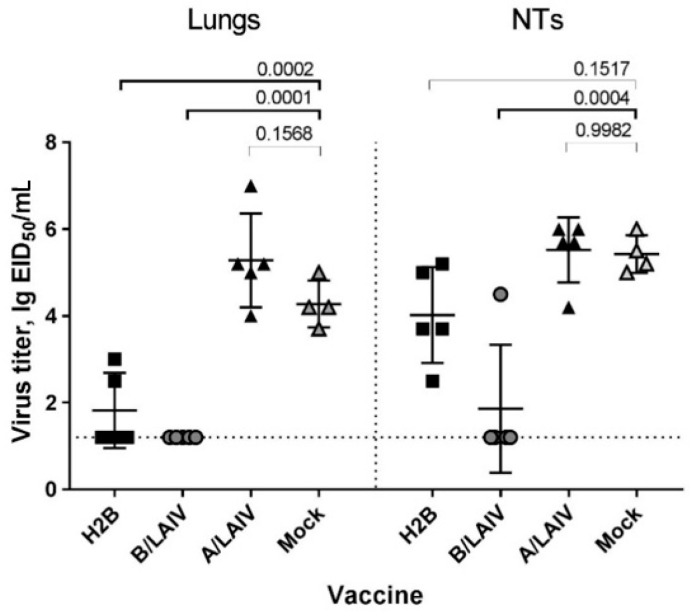
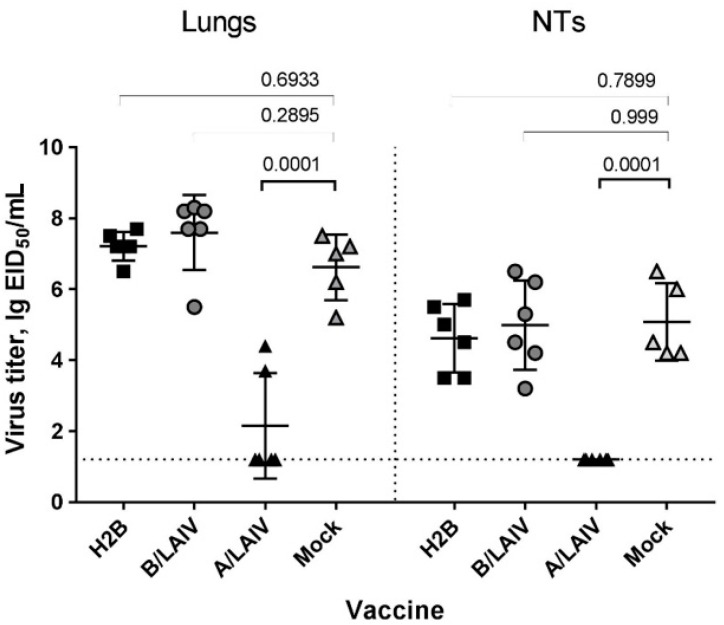
Influenza A and B viruses cause significant morbidity and mortality worldwide. Current influenza vaccines are composed of three or four strains: A/H1N1, A/H3N2, and B (Victoria and Yamagata lineages). It is of great interest if immunization against both type A and B influenza viruses can be combined in a single vaccine strain, thus reducing the cost of vaccine production and the possibility of strain interference within the multicomponent vaccine. In the current study, we developed an experimental live cold-adapted influenza intertype reassortant (influenza A and B) vaccine on the live attenuated influenza vaccine (LAIV) A/Leningrad/134/17/57 backbone. Hemagglutinin (HA) and neuraminidase (NA) functional domains were inherited from the influenza B/Brisbane/60/2008 strain, whereas their packaging signals were substituted with appropriate fragments of influenza A virus genes. The recombinant A/B virus efficiently replicated in eggs and Madin-Darby Canine Kidney (MDCK) cells under optimal conditions, temperature-sensitive phenotype was maintained, and its antigenic properties matched the influenza B parental virus. The chimeric vaccine was attenuated in mice: after intranasal immunization, viral replication was seen only in nasal turbinates but not in the lungs. Immunological studies demonstrated the induction of IgG antibody responses against the influenza A and B virus, whereas hemagglutination inhibition (HAI) and neutralizing antibodies were detected only against the influenza B virus, resulting in significant protection of immunized animals against influenza B virus challenge. IFNγ-secreting CD8 effector memory T cells (CD44+CD62L-) were detected in mouse splenocytes after stimulation with the specific influenza A peptide (NP366); however, the T-cell response was not sufficient to protect animals against infection with a high-dose mouse-adapted A/California/07/2009 (H1N1pdm09) virus, most probably due to the mismatch of key T-cell epitopes of the H1N1 virus and the LAIV backbone. Overall, generation of the chimeric A/B LAIV virus on a licensed LAIV backbone demonstrated prospects for the development of safe and efficacious vaccine candidates that afford combined protection against both type A and type B influenza viruses; however, further optimization of the T-cell epitope content within the LAIV backbone may be required.
Keywords: LAIV; chimeric hemagglutinin; genome packaging; influenza B virus; influenza virus; live attenuated influenza vaccine; reassortment; universal influenza vaccine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Safety, Immunogenicity, and Protective Efficacy of an H5N1 Chimeric Cold-Adapted Attenuated Virus Vaccine in a Mouse Model.Viruses. 2021 Dec 3;13(12):2420. doi: 10.3390/v13122420. Viruses. 2021. PMID: 34960689 Free PMC article.
-
Sequential Immunization With Live-Attenuated Chimeric Hemagglutinin-Based Vaccines Confers Heterosubtypic Immunity Against Influenza A Viruses in a Preclinical Ferret Model.Front Immunol. 2019 Apr 10;10:756. doi: 10.3389/fimmu.2019.00756. eCollection 2019. Front Immunol. 2019. PMID: 31105689 Free PMC article.
-
Safety and immunogenicity in man of a cell culture derived trivalent live attenuated seasonal influenza vaccine: a Phase I dose escalating study in healthy volunteers.Vaccine. 2014 Sep 3;32(39):5118-24. doi: 10.1016/j.vaccine.2014.05.030. Epub 2014 May 23. Vaccine. 2014. PMID: 24858566 Clinical Trial.
-
Influenza virus vaccine live intranasal--MedImmune vaccines: CAIV-T, influenza vaccine live intranasal.Drugs R D. 2003;4(5):312-9. doi: 10.2165/00126839-200304050-00007. Drugs R D. 2003. PMID: 12952502 Review.
-
Live Attenuated Reassortant Vaccines Based on A/Leningrad/134/17/57 Master Donor Virus Against H5 Avian Influenza.Open Microbiol J. 2017 Nov 30;11:316-329. doi: 10.2174/1874285801711010316. eCollection 2017. Open Microbiol J. 2017. PMID: 29290844 Free PMC article. Review.
Cited by
-
Safety, Immunogenicity, and Protective Efficacy of an H5N1 Chimeric Cold-Adapted Attenuated Virus Vaccine in a Mouse Model.Viruses. 2021 Dec 3;13(12):2420. doi: 10.3390/v13122420. Viruses. 2021. PMID: 34960689 Free PMC article.
-
Influenza B: Prospects for the Development of Cross-Protective Vaccines.Viruses. 2022 Jun 17;14(6):1323. doi: 10.3390/v14061323. Viruses. 2022. PMID: 35746794 Free PMC article. Review.
-
Clade 2.3.4.4 H5 chimeric cold-adapted attenuated influenza vaccines induced cross-reactive protection in mice and ferrets.J Virol. 2023 Nov 30;97(11):e0110123. doi: 10.1128/jvi.01101-23. Epub 2023 Nov 2. J Virol. 2023. PMID: 37916835 Free PMC article.
References
-
- Hoft D.F., Lottenbach K.R., Blazevic A., Turan A., Blevins T.P., Pacatte T.P., Yu Y., Mitchell M.C., Hoft S.G., Belshe R.B. Comparisons of the Humoral and Cellular Immune Responses Induced by Live Attenuated Influenza Vaccine and Inactivated Influenza Vaccine in Adults. Clin. Vaccine Immunol. 2016;24 doi: 10.1128/CVI.00414-16. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
