

Permissive selection followed by affinity-based proliferation of GC light zone B cells dictates cell fate and ensures clonal breadth
- PMID: 33419925
- PMCID: PMC7812803
- DOI: 10.1073/pnas.2016425118
Permissive selection followed by affinity-based proliferation of GC light zone B cells dictates cell fate and ensures clonal breadth
Abstract
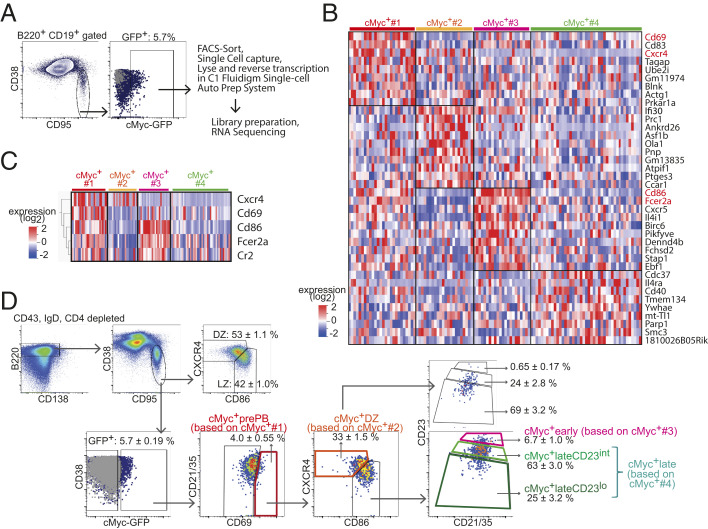
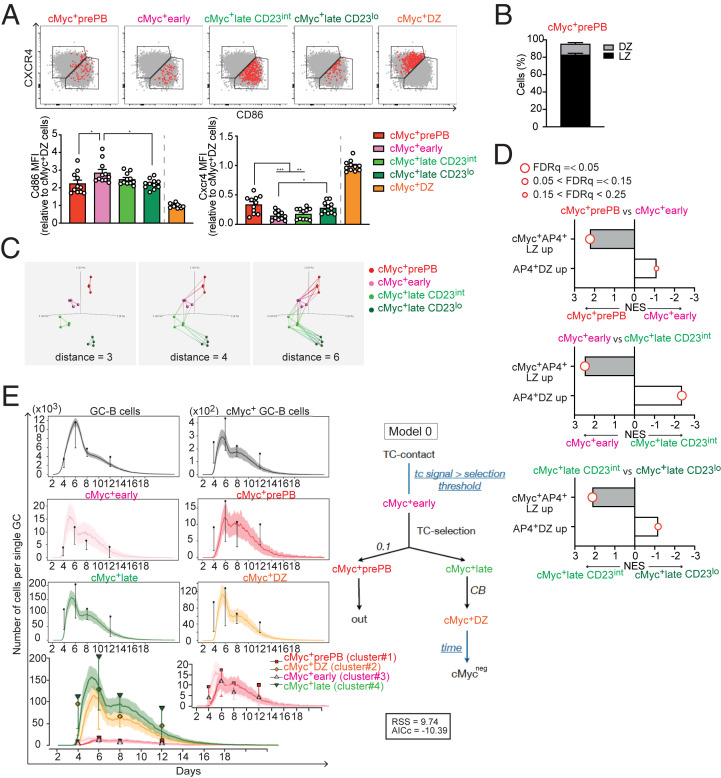
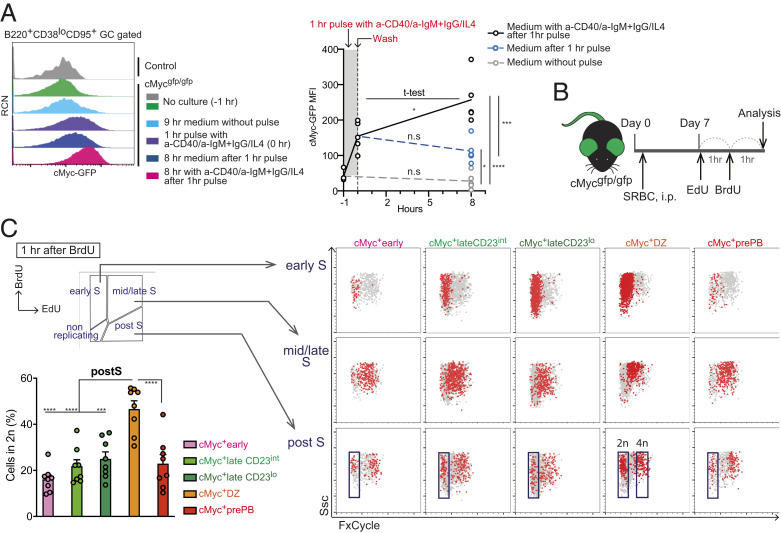
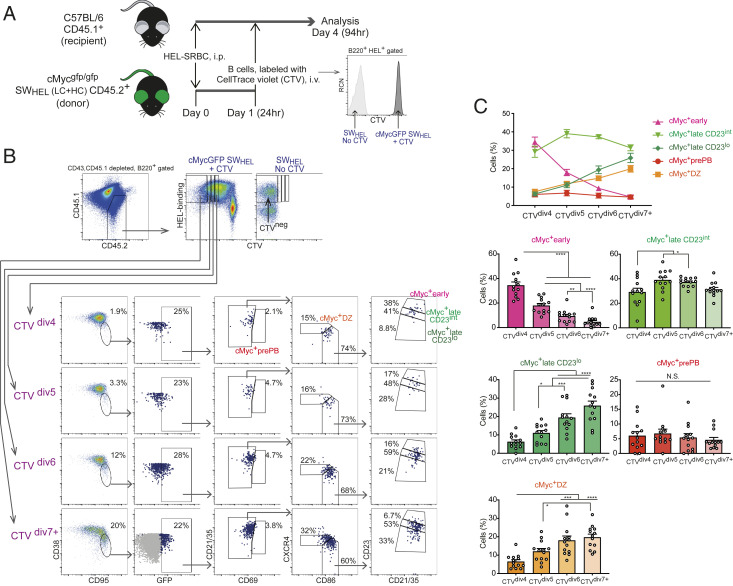
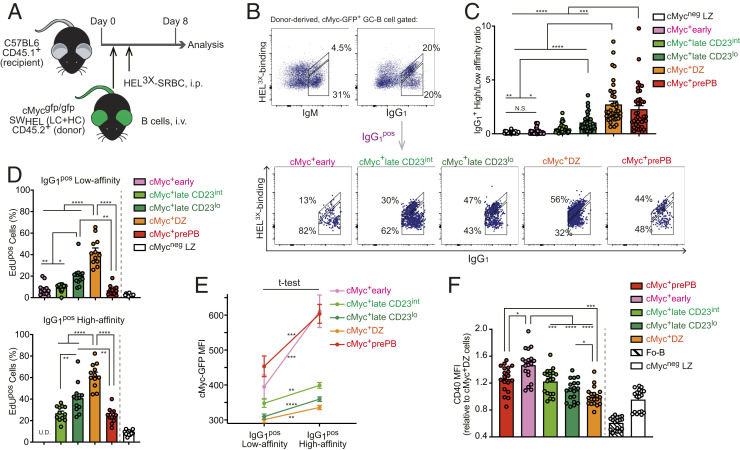
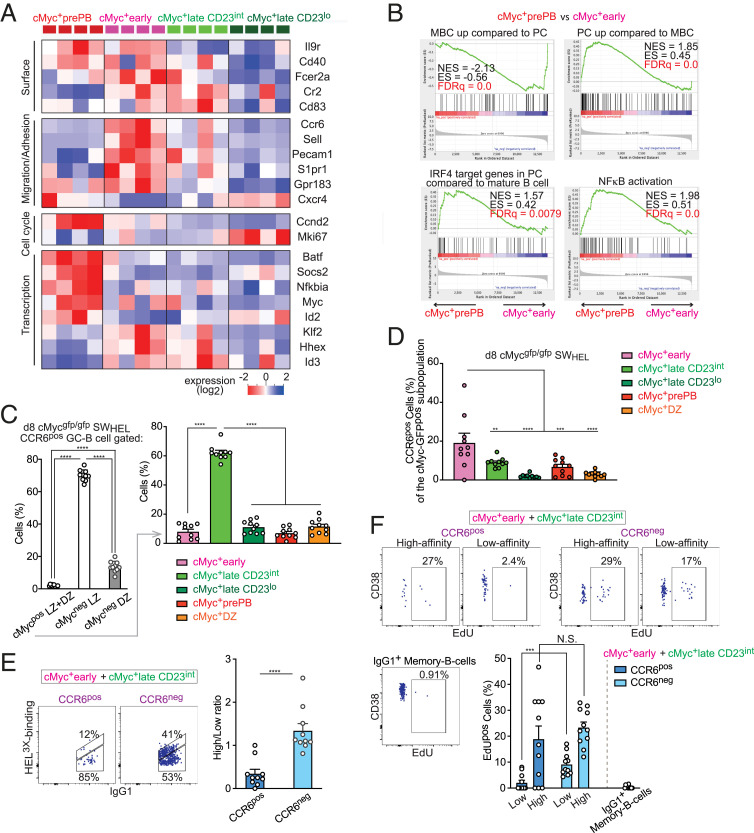
Affinity maturation depends on how efficiently germinal centers (GCs) positively select B cells in the light zone (LZ). Positively selected GC B cells recirculate between LZs and dark zones (DZs) and ultimately differentiate into plasmablasts (PBs) and memory B cells (MBCs). Current understanding of the GC reaction presumes that cMyc-dependent positive selection of LZ B cells is a competitive affinity-dependent process; however, this cannot explain the production of GC-derived lower-affinity MBCs or retention of GC B cells with varied affinities. Here, by combining single-cell/bulk RNA sequencing and flow cytometry, we identified and characterized temporally and functionally distinct positively selected cMyc+ GC B cell subpopulations. cMyc+ LZ B cell subpopulations enriched with either higher- or lower-affinity cells diverged soon after permissive positive selection. The former subpopulation contained PB precursors, whereas the latter comprised less proliferative MBC precursors and future DZ entrants. The overall affinity of future DZ entrants was enhanced in the LZ through preferential proliferation of higher-affinity cells. Concurrently, lower-affinity cells were retained in GCs and protected from apoptosis. These findings redefine positive selection as a dynamic process generating three distinct B cell fates and elucidate how positive selection ensures clonal diversity for broad protection.
Keywords: GC B cells; affinity maturation; clonal diversity; memory B cells; positive selection.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Positive Selection in the Light Zone of Germinal Centers.Front Immunol. 2021 Mar 31;12:661678. doi: 10.3389/fimmu.2021.661678. eCollection 2021. Front Immunol. 2021. PMID: 33868314 Free PMC article. Review.
-
Germinal Center B Cells Replace Their Antigen Receptors in Dark Zones and Fail Light Zone Entry when Immunoglobulin Gene Mutations are Damaging.Immunity. 2018 Sep 18;49(3):477-489.e7. doi: 10.1016/j.immuni.2018.08.025. Immunity. 2018. PMID: 30231983 Free PMC article.
-
Germinal Center Selection and Affinity Maturation Require Dynamic Regulation of mTORC1 Kinase.Immunity. 2017 Jun 20;46(6):1045-1058.e6. doi: 10.1016/j.immuni.2017.06.005. Immunity. 2017. PMID: 28636954 Free PMC article.
-
Cbl Ubiquitin Ligases Control B Cell Exit from the Germinal-Center Reaction.Immunity. 2018 Mar 20;48(3):530-541.e6. doi: 10.1016/j.immuni.2018.03.006. Immunity. 2018. PMID: 29562201
-
Heterogeneity of germinal center B cells: New insights from single-cell studies.Eur J Immunol. 2021 Nov;51(11):2555-2567. doi: 10.1002/eji.202149235. Epub 2021 Aug 8. Eur J Immunol. 2021. PMID: 34324199 Review.
Cited by
-
Epidermal stem cells participate in the repair of scalds via Nanog and Myc regulation.Mol Med Rep. 2022 Dec;26(6):364. doi: 10.3892/mmr.2022.12881. Epub 2022 Oct 25. Mol Med Rep. 2022. PMID: 36281911 Free PMC article.
-
B cells require DOCK8 to elicit and integrate T cell help when antigen is limiting.Sci Immunol. 2024 Aug 9;9(98):eadd4874. doi: 10.1126/sciimmunol.add4874. Epub 2024 Aug 9. Sci Immunol. 2024. PMID: 39121196 Free PMC article.
-
Investigating the Mechanism of Germinal Center Shutdown.Front Immunol. 2022 Jul 14;13:922318. doi: 10.3389/fimmu.2022.922318. eCollection 2022. Front Immunol. 2022. PMID: 35911680 Free PMC article.
-
Competition for refueling rather than cyclic reentry initiation evident in germinal centers.Sci Immunol. 2022 Mar 11;7(69):eabm0775. doi: 10.1126/sciimmunol.abm0775. Epub 2022 Mar 11. Sci Immunol. 2022. PMID: 35275753 Free PMC article.
-
Bet-hedging in innate and adaptive immune systems.Evol Med Public Health. 2022 May 24;10(1):256-265. doi: 10.1093/emph/eoac021. eCollection 2022. Evol Med Public Health. 2022. PMID: 35712085 Free PMC article.
References
-
- Berek C., Berger A., Apel M., Maturation of the immune response in germinal centers. Cell 67, 1121–1129 (1991). - PubMed
-
- Jacob J., Kelsoe G., Rajewsky K., Weiss U., Intraclonal generation of antibody mutants in germinal centres. Nature 354, 389–392 (1991). - PubMed
-
- Kepler T. B., Perelson A. S., Cyclic re-entry of germinal center B cells and the efficiency of affinity maturation. Immunol. Today 14, 412–415 (1993). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
