

Single-Cell Sequencing of Brain Cell Transcriptomes and Epigenomes
- PMID: 33412093
- PMCID: PMC7808568
- DOI: 10.1016/j.neuron.2020.12.010
Single-Cell Sequencing of Brain Cell Transcriptomes and Epigenomes
Abstract
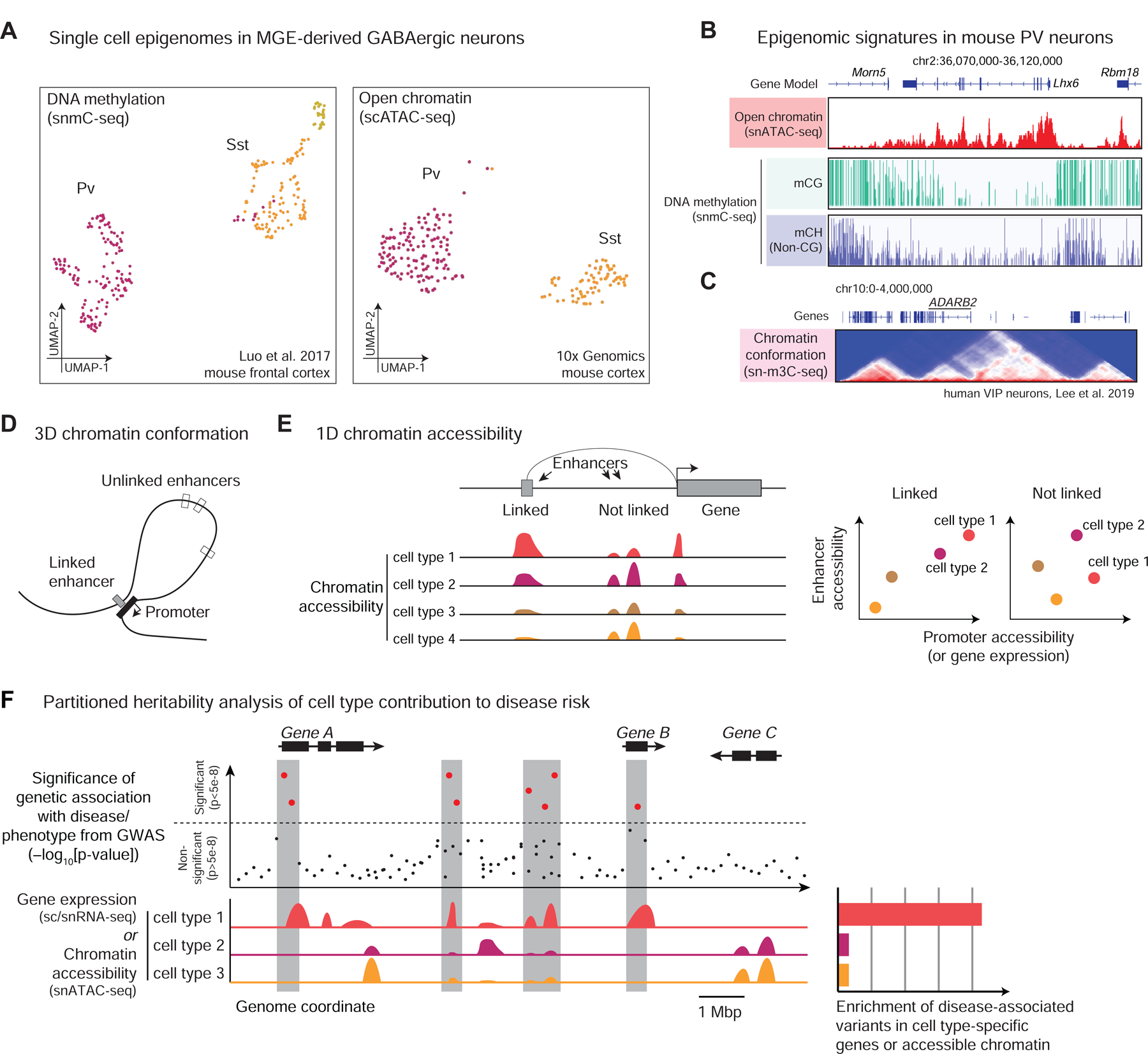
Single-cell sequencing technologies, including transcriptomic and epigenomic assays, are transforming our understanding of the cellular building blocks of neural circuits. By directly measuring multiple molecular signatures in thousands to millions of individual cells, single-cell sequencing methods can comprehensively characterize the diversity of brain cell types. These measurements uncover gene regulatory mechanisms that shape cellular identity and provide insight into developmental and evolutionary relationships between brain cell populations. Single-cell sequencing data can aid the design of tools for targeted functional studies of brain circuit components, linking molecular signatures with anatomy, connectivity, morphology, and physiology. Here, we discuss the fundamental principles of single-cell transcriptome and epigenome sequencing, integrative computational analysis of the data, and key applications in neuroscience.
Keywords: ATAC-seq; DNA methylation; cell state; cell type; epigenome; multi-omics; open chromatin; single-cell sequencing; spatial transcriptomics; transcriptome.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures


Similar articles
-
EpiScanpy: integrated single-cell epigenomic analysis.Nat Commun. 2021 Sep 1;12(1):5228. doi: 10.1038/s41467-021-25131-3. Nat Commun. 2021. PMID: 34471111 Free PMC article.
-
Single-Cell Sequencing Methodologies: From Transcriptome to Multi-Dimensional Measurement.Small Methods. 2021 Jun;5(6):e2100111. doi: 10.1002/smtd.202100111. Epub 2021 Apr 17. Small Methods. 2021. PMID: 34927917 Review.
-
Single-Cell Analysis of the Transcriptome and Epigenome.Methods Mol Biol. 2022;2399:21-60. doi: 10.1007/978-1-0716-1831-8_3. Methods Mol Biol. 2022. PMID: 35604552 Free PMC article.
-
Simultaneous Profiling of mRNA Transcriptome and DNA Methylome from a Single Cell.Methods Mol Biol. 2019;1979:363-377. doi: 10.1007/978-1-4939-9240-9_21. Methods Mol Biol. 2019. PMID: 31028648
-
Deciphering Brain Complexity Using Single-cell Sequencing.Genomics Proteomics Bioinformatics. 2019 Aug;17(4):344-366. doi: 10.1016/j.gpb.2018.07.007. Epub 2019 Oct 3. Genomics Proteomics Bioinformatics. 2019. PMID: 31586689 Free PMC article. Review.
Cited by
-
Recent Major Transcriptomics and Epitranscriptomics Contributions toward Personalized and Precision Medicine.J Pers Med. 2022 Feb 1;12(2):199. doi: 10.3390/jpm12020199. J Pers Med. 2022. PMID: 35207687 Free PMC article. Review.
-
Dopamine Neuron Diversity: Recent Advances and Current Challenges in Human Stem Cell Models and Single Cell Sequencing.Cells. 2021 Jun 1;10(6):1366. doi: 10.3390/cells10061366. Cells. 2021. PMID: 34206038 Free PMC article. Review.
-
Evolution of glutamatergic signaling and synapses.Neuropharmacology. 2021 Nov 1;199:108740. doi: 10.1016/j.neuropharm.2021.108740. Epub 2021 Jul 31. Neuropharmacology. 2021. PMID: 34343611 Free PMC article. Review.
-
A transcriptomic and epigenomic cell atlas of the mouse primary motor cortex.Nature. 2021 Oct;598(7879):103-110. doi: 10.1038/s41586-021-03500-8. Epub 2021 Oct 6. Nature. 2021. PMID: 34616066 Free PMC article.
-
Celloscope: a probabilistic model for marker-gene-driven cell type deconvolution in spatial transcriptomics data.Genome Biol. 2023 May 17;24(1):120. doi: 10.1186/s13059-023-02951-8. Genome Biol. 2023. PMID: 37198601 Free PMC article.
References
-
- Arendt D, Musser JM, Baker CVH, Bergman A, Cepko C, Erwin DH, Pavlicev M, Schlosser G, Widder S, Laubichler MD, et al. (2016). The origin and evolution of cell types. Nat. Rev. Genet 17, 744–757. - PubMed
-
- Azevedo FAC, Carvalho LRB, Grinberg LT, Farfel JM, Ferretti REL, Leite REP, Jacob Filho W, Lent R, and Herculano-Houzel S (2009). Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol 513, 532–541. - PubMed
-
- Bakken TE, Jorstad NL, Hu Q, Lake BB, and Tian W (2020). Evolution of cellular diversity in primary motor cortex of human, marmoset monkey, and mouse. bioRxiv.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
