

The functional organization of axonal mRNA transport and translation
- PMID: 33288912
- PMCID: PMC8161363
- DOI: 10.1038/s41583-020-00407-7
The functional organization of axonal mRNA transport and translation
Abstract
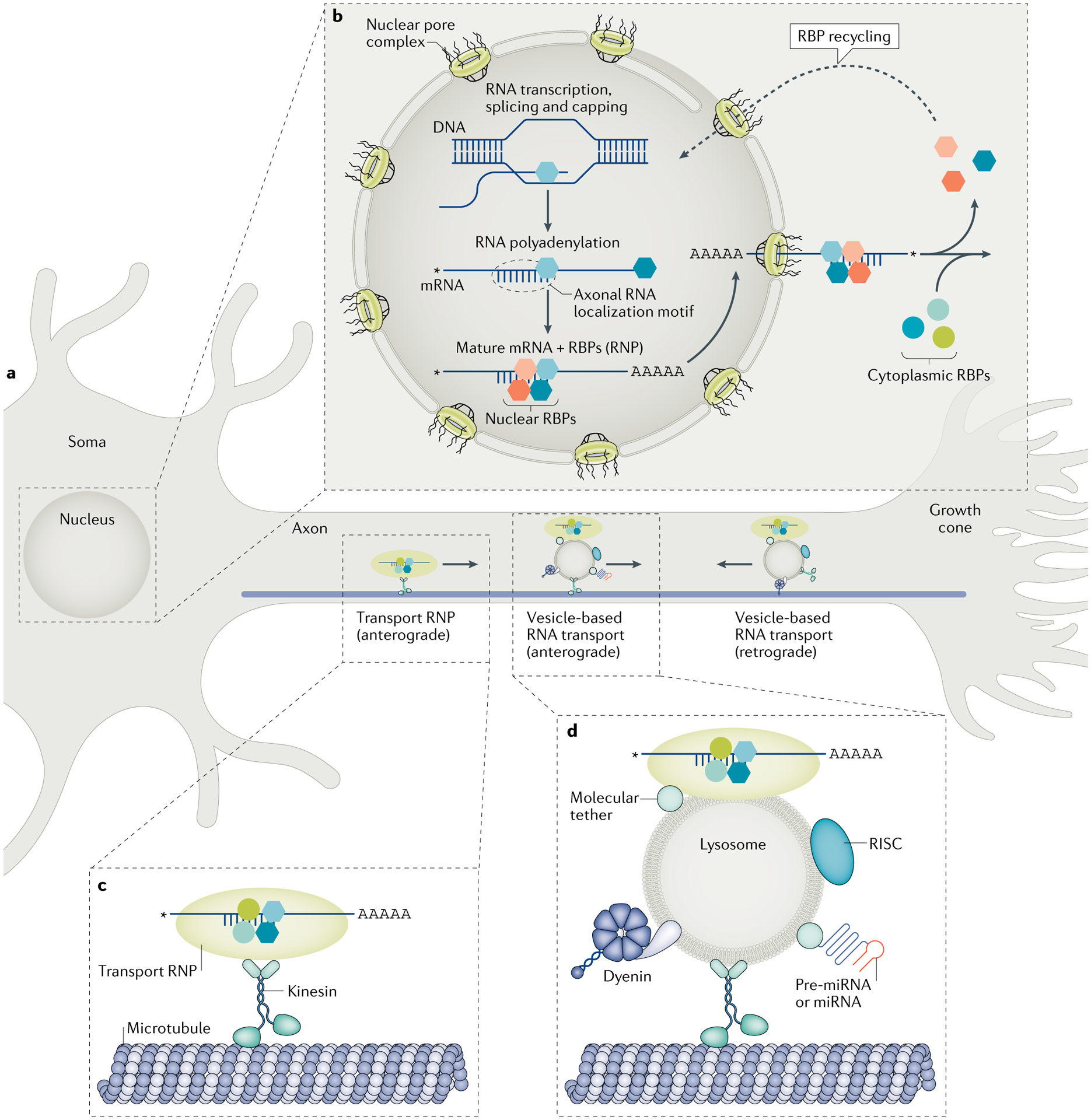
Axons extend for tremendously long distances from the neuronal soma and make use of localized mRNA translation to rapidly respond to different extracellular stimuli and physiological states. The locally synthesized proteins support many different functions in both developing and mature axons, raising questions about the mechanisms by which local translation is organized to ensure the appropriate responses to specific stimuli. Publications over the past few years have uncovered new mechanisms for regulating the axonal transport and localized translation of mRNAs, with several of these pathways converging on the regulation of cohorts of functionally related mRNAs - known as RNA regulons - that drive axon growth, axon guidance, injury responses, axon survival and even axonal mitochondrial function. Recent advances point to these different regulatory pathways as organizing platforms that allow the axon's proteome to be modulated to meet its physiological needs.
Conflict of interest statement
Competing interests
J,L.T. and P.K.S. have a US Patent for G3BP1 as a target for accelerating axon regeneration (US Patent 10,668,128). A.N.K., P.K.S. and J.L.T. have applied for a US patent for G3BP1 as a target for preventing neurodegeneration.
Figures



Similar articles
-
Regulation of mRNA transport and translation in axons.Results Probl Cell Differ. 2009;48:193-224. doi: 10.1007/400_2009_16. Results Probl Cell Differ. 2009. PMID: 19582411 Review.
-
Molecular control of local translation in axon development and maintenance.Curr Opin Neurobiol. 2018 Aug;51:86-94. doi: 10.1016/j.conb.2018.02.025. Epub 2018 Mar 14. Curr Opin Neurobiol. 2018. PMID: 29549711 Review.
-
Dynamics of axonal mRNA transport and implications for peripheral nerve regeneration.Exp Neurol. 2010 May;223(1):19-27. doi: 10.1016/j.expneurol.2009.08.011. Epub 2009 Aug 20. Exp Neurol. 2010. PMID: 19699200 Free PMC article. Review.
-
Local protein synthesis in neuronal axons: why and how we study.BMB Rep. 2015 Mar;48(3):139-46. doi: 10.5483/bmbrep.2015.48.3.010. BMB Rep. 2015. PMID: 25644635 Free PMC article. Review.
-
Axonal transport of neural membrane protein 35 mRNA increases axon growth.J Cell Sci. 2013 Jan 1;126(Pt 1):90-102. doi: 10.1242/jcs.107268. Epub 2012 Oct 24. J Cell Sci. 2013. PMID: 23097042 Free PMC article.
Cited by
-
hnRNP R promotes O-GlcNAcylation of eIF4G and facilitates axonal protein synthesis.Nat Commun. 2024 Aug 28;15(1):7430. doi: 10.1038/s41467-024-51678-y. Nat Commun. 2024. PMID: 39198412 Free PMC article.
-
Cells of the Central Nervous System: An Overview of Their Structure and Function.Adv Neurobiol. 2023;29:41-64. doi: 10.1007/978-3-031-12390-0_2. Adv Neurobiol. 2023. PMID: 36255671
-
Neurofilament Light Regulates Axon Caliber, Synaptic Activity, and Organelle Trafficking in Cultured Human Motor Neurons.Front Cell Dev Biol. 2022 Feb 14;9:820105. doi: 10.3389/fcell.2021.820105. eCollection 2021. Front Cell Dev Biol. 2022. PMID: 35237613 Free PMC article.
-
Distinct small non-coding RNA landscape in the axons and released extracellular vesicles of developing primary cortical neurons and the axoplasm of adult nerves.RNA Biol. 2021 Nov 12;18(sup2):832-855. doi: 10.1080/15476286.2021.2000792. Epub 2021 Dec 9. RNA Biol. 2021. PMID: 34882524 Free PMC article.
-
MicroRNAs 21 and 199a-3p Regulate Axon Growth Potential through Modulation of Pten and mTor mRNAs.eNeuro. 2021 Aug 11;8(4):ENEURO.0155-21.2021. doi: 10.1523/ENEURO.0155-21.2021. Print 2021 Jul-Aug. eNeuro. 2021. PMID: 34326064 Free PMC article.
References
-
- Steward O & Schuman EM Protein synthesis at synaptic sites on dendrites. Annu. Rev. Neurosci 24, 299–325 (2001). - PubMed
-
- Kosik KS Life at low copy number: how dendrites manage with so few mRNAs. Neuron 92, 1168–1180 (2016). - PubMed
-
- Tobias G & Koenig E Axonal protein synthesizing activity during the early outgrowth period following neurotomy. Exp. Neurol 49, 221–234 (1975). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
