

Defined Intestinal Regions Are Drained by Specific Lymph Nodes That Mount Distinct Th1 and Th2 Responses Against Schistosoma mansoni Eggs
- PMID: 33193437
- PMCID: PMC7644866
- DOI: 10.3389/fimmu.2020.592325
Defined Intestinal Regions Are Drained by Specific Lymph Nodes That Mount Distinct Th1 and Th2 Responses Against Schistosoma mansoni Eggs
Abstract
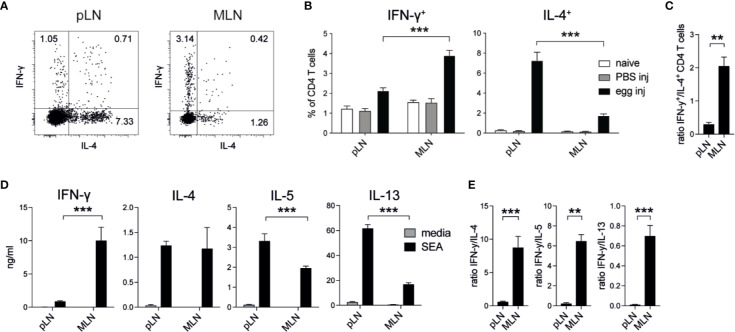
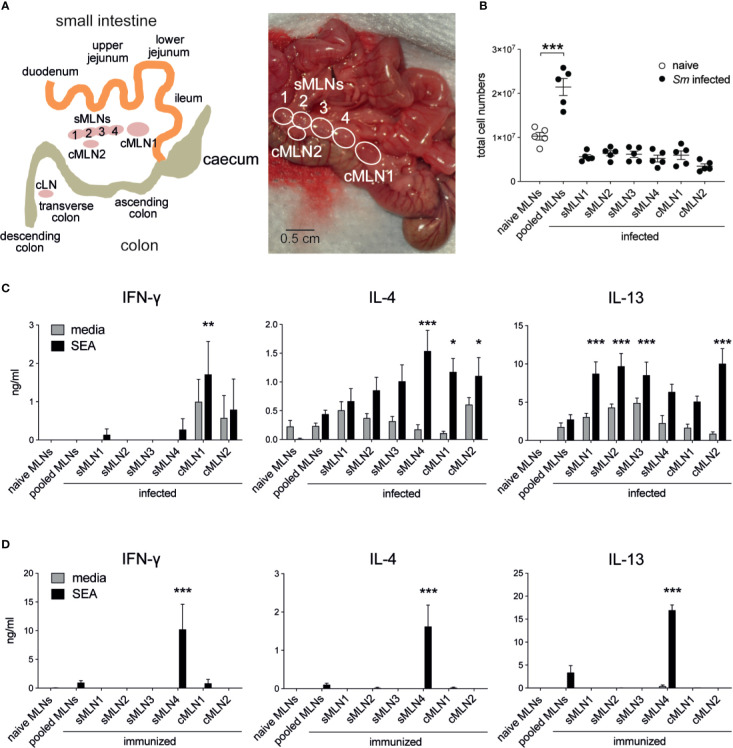
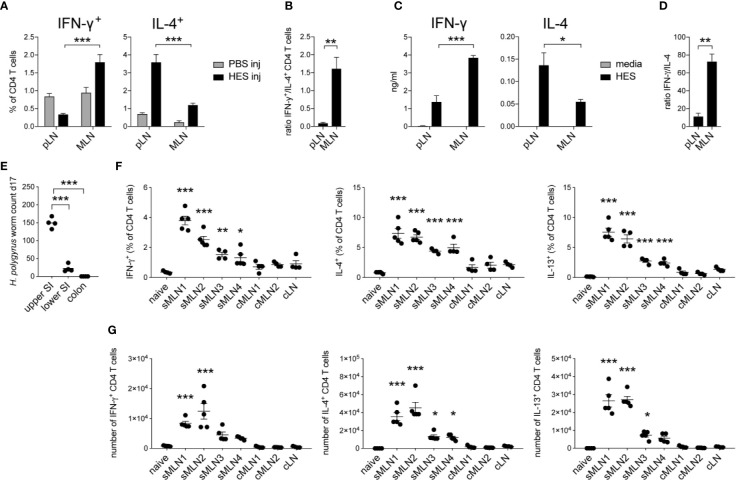
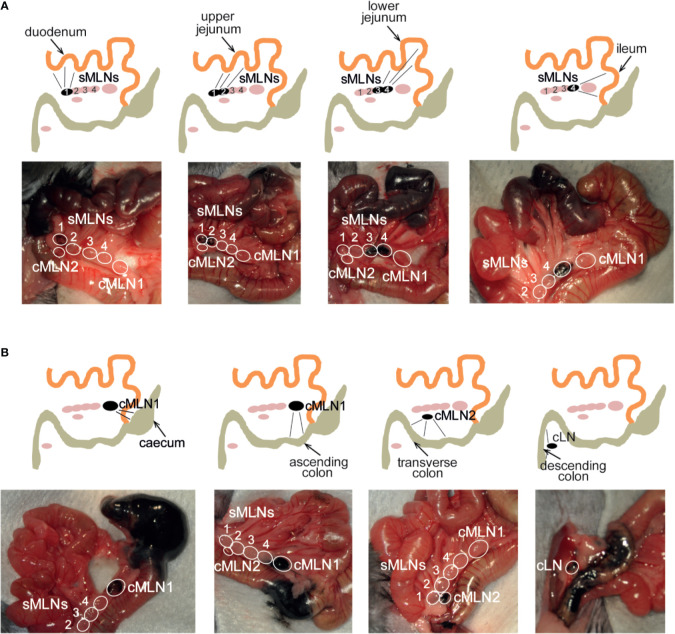
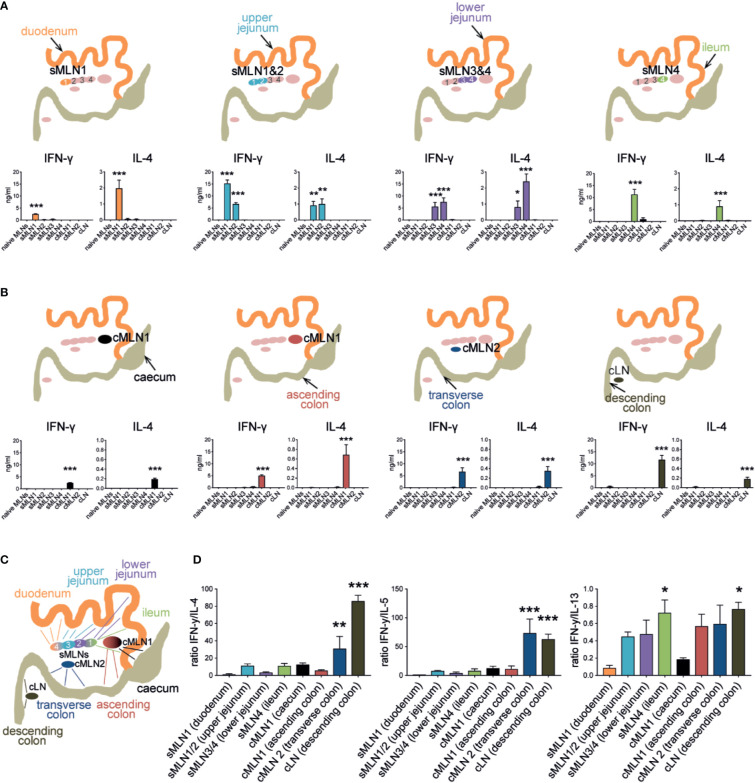
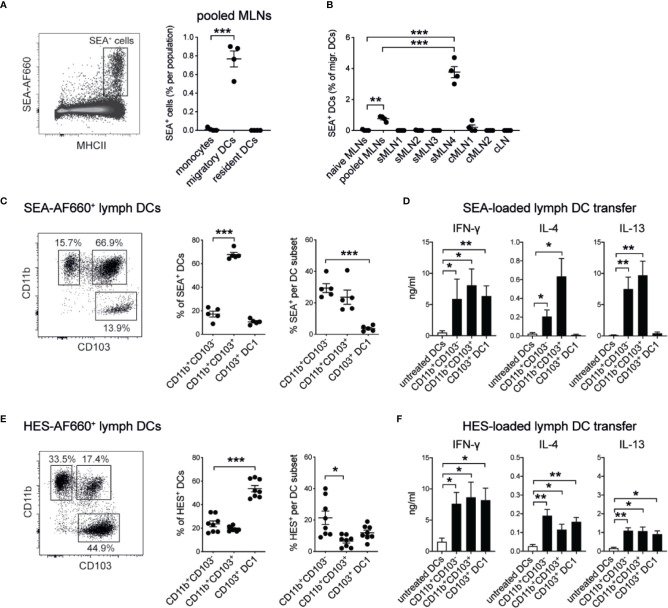
The balance of type 1 and type 2 immune responses plays a crucial role in anti-helminth immunity and can either support chronic infection or drive type 2 mediated expulsion of the parasite. Helminth antigens and secreted molecules directly influence this balance and induce a favorable immunological environment for the parasite's survival. However, less is known if the site of infection also influences the balance of type 1 and type 2 immunity. Here, we report that tissue-specific immune responses are mounted against helminth antigens, which elicited strong IL-4 responses when injected into the skin, while the same antigen, delivered into the intestinal subserosa, induced increased IFN-γ and reduced Th2 responses. Immune responses in individual mesenteric lymph nodes that drain defined regions of the intestine furthermore displayed a site-specific pattern of type 1 and type 2 immunity after Schistosoma mansoni or Heligmosomoides polygyrus infection. S. mansoni egg-specific Th2 responses were detectable in all mesenteric lymph nodes but Th1 responses were only present in those draining the colon, while H. polygyrus infection elicited mixed Th1 and Th2 responses in the lymph nodes associated with the site of infection. Similar site-specific type 1 and type 2 immune responses were observed in the draining lymph nodes after the controlled delivery of S. mansoni eggs into different segments of the small and large intestine using microsurgical techniques. Different subsets of intestinal dendritic cells were hereby responsible for the uptake and priming of Th1 and Th2 responses against helminth antigens. Migratory CD11b+CD103- and especially CD11b+CD103+ DC2s transported S. mansoni egg antigens to the draining lymph nodes to induce Th1 and Th2 responses, while CD103+ DC1s induced only IFN-γ responses. In contrast, H. polygyrus antigens were predominantly transported by CD11b+CD103- DC2s and CD103+ DC1s and all DC subsets induced similar Th1 but weaker Th2 responses, compared to S. mansoni egg antigens. The development of adaptive anti-helminth immune responses is therefore influenced by the antigen itself, the uptake and priming characteristics of antigen-positive dendritic cell subsets and the site of infection, which shape the level of Th1 and Th2 responses in order to create a favorable immunological environment for the parasite.
Keywords: Heligmosomoides polygyrus bakeri; Schistosoma mansoni; dendritic cells; helminth antigen; mesenteric lymph node; microsurgery; mucosal immunology; th1/th2 balance.
Copyright © 2020 Mayer, Brown, MacDonald and Milling.
Figures
Similar articles
-
Plasmacytoid Dendritic Cells Facilitate Th Cell Cytokine Responses throughout Schistosoma mansoni Infection.Immunohorizons. 2021 Aug 30;5(8):721-732. doi: 10.4049/immunohorizons.2100071. Immunohorizons. 2021. PMID: 34462311 Free PMC article.
-
T-helper type-1-dominated lymph node responses induced in C57BL/6 mice by optimally irradiated cercariae of Schistosoma mansoni are down-regulated after challenge infection.Immunology. 1995 Feb;84(2):310-6. Immunology. 1995. PMID: 7751008 Free PMC article.
-
In infection with Schistosoma mansoni, B cells are required for T helper type 2 cell responses but not for granuloma formation.J Immunol. 1997 May 15;158(10):4832-7. J Immunol. 1997. PMID: 9144498
-
A double edged sword: Schistosoma mansoni Sm29 regulates both Th1 and Th2 responses in inflammatory mucosal diseases.Mucosal Immunol. 2016 Nov;9(6):1366-1371. doi: 10.1038/mi.2016.69. Epub 2016 Aug 24. Mucosal Immunol. 2016. PMID: 27554296 Review. No abstract available.
-
Th2 response polarization during infection with the helminth parasite Schistosoma mansoni.Immunol Rev. 2004 Oct;201:117-26. doi: 10.1111/j.0105-2896.2004.00187.x. Immunol Rev. 2004. PMID: 15361236 Review.
Cited by
-
Conventional Type 1 Dendritic Cells in Intestinal Immune Homeostasis.Front Immunol. 2022 May 27;13:857954. doi: 10.3389/fimmu.2022.857954. eCollection 2022. Front Immunol. 2022. PMID: 35693801 Free PMC article. Review.
-
Immune Enhancement of Nanoparticle-Encapsulated Ginseng Stem-Leaf Saponins on Porcine Epidemic Diarrhea Virus Vaccine in Mice.Vaccines (Basel). 2022 Oct 27;10(11):1810. doi: 10.3390/vaccines10111810. Vaccines (Basel). 2022. PMID: 36366319 Free PMC article.
-
Basophils control T cell priming through soluble mediators rather than antigen presentation.Front Immunol. 2023 Feb 9;13:1032379. doi: 10.3389/fimmu.2022.1032379. eCollection 2022. Front Immunol. 2023. PMID: 36846020 Free PMC article. Review.
-
Anti-Trichuris mucosal responses are maintained during H. bakeri co-infection despite impaired parasite expulsion.Parasite Immunol. 2022 Aug;44(8):e12936. doi: 10.1111/pim.12936. Epub 2022 May 31. Parasite Immunol. 2022. PMID: 35586956 Free PMC article.
-
Antigen-level resolution of commensal-specific B cell responses can be enabled by phage display screening coupled with B cell tetramers.Immunity. 2024 Jun 11;57(6):1428-1441.e8. doi: 10.1016/j.immuni.2024.04.014. Epub 2024 May 8. Immunity. 2024. PMID: 38723638
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials