

Meiosis reveals the early steps in the evolution of a neo-XY sex chromosome pair in the African pygmy mouse Mus minutoides
- PMID: 33180767
- PMCID: PMC7685469
- DOI: 10.1371/journal.pgen.1008959
Meiosis reveals the early steps in the evolution of a neo-XY sex chromosome pair in the African pygmy mouse Mus minutoides
Abstract
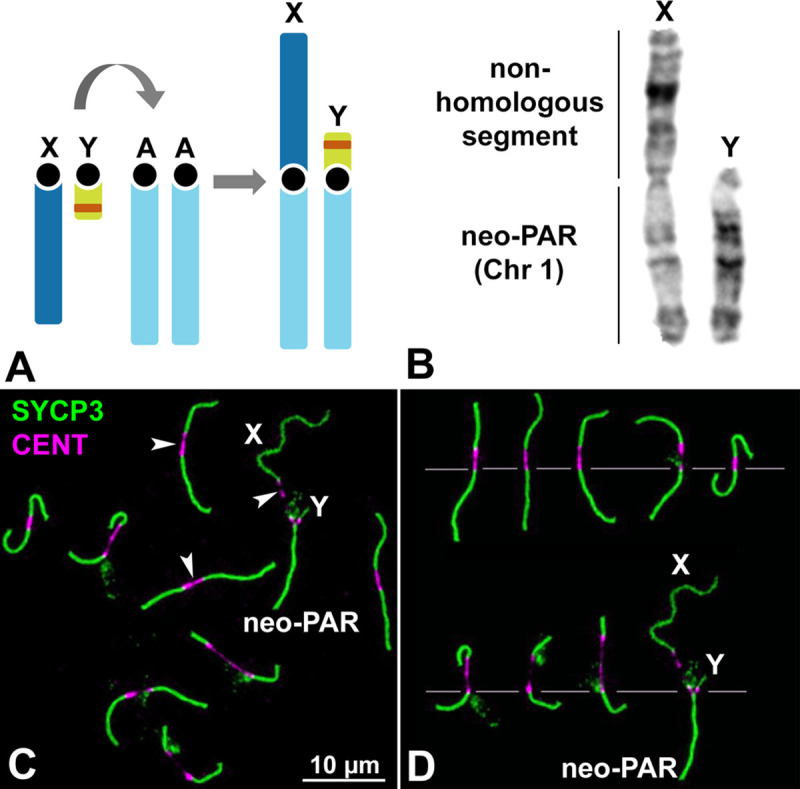
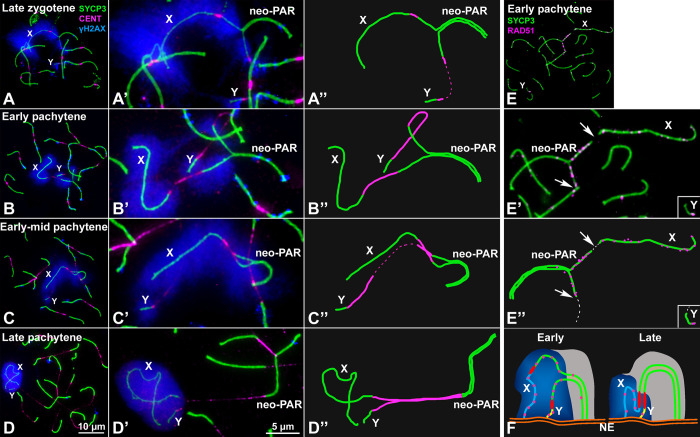
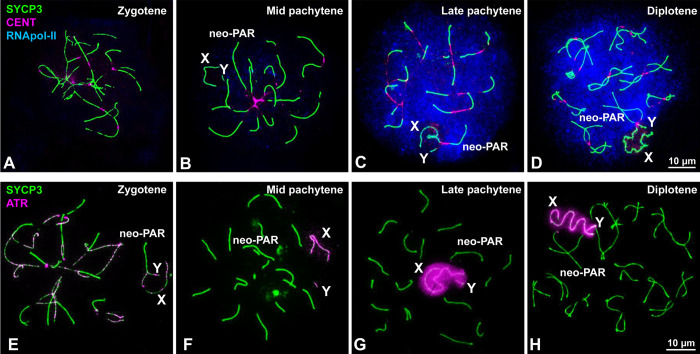
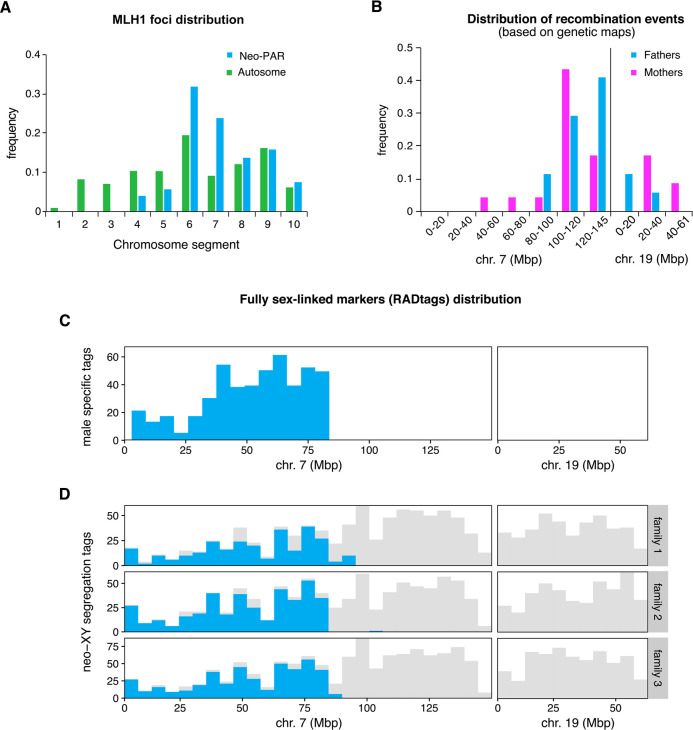
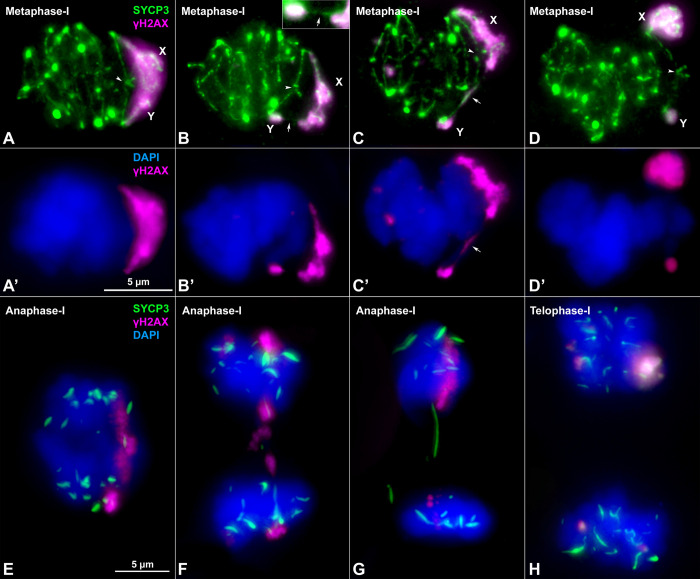
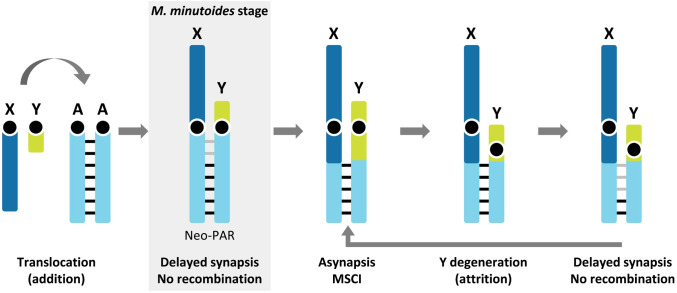
Sex chromosomes of eutherian mammals are highly different in size and gene content, and share only a small region of homology (pseudoautosomal region, PAR). They are thought to have evolved through an addition-attrition cycle involving the addition of autosomal segments to sex chromosomes and their subsequent differentiation. The events that drive this process are difficult to investigate because sex chromosomes in almost all mammals are at a very advanced stage of differentiation. Here, we have taken advantage of a recent translocation of an autosome to both sex chromosomes in the African pygmy mouse Mus minutoides, which has restored a large segment of homology (neo-PAR). By studying meiotic sex chromosome behavior and identifying fully sex-linked genetic markers in the neo-PAR, we demonstrate that this region shows unequivocal signs of early sex-differentiation. First, synapsis and resolution of DNA damage intermediates are delayed in the neo-PAR during meiosis. Second, recombination is suppressed or largely reduced in a large portion of the neo-PAR. However, the inactivation process that characterizes sex chromosomes during meiosis does not extend to this region. Finally, the sex chromosomes show a dual mechanism of association at metaphase-I that involves the formation of a chiasma in the neo-PAR and the preservation of an ancestral achiasmate mode of association in the non-homologous segments. We show that the study of meiosis is crucial to apprehend the onset of sex chromosome differentiation, as it introduces structural and functional constrains to sex chromosome evolution. Synapsis and DNA repair dynamics are the first processes affected in the incipient differentiation of X and Y chromosomes, and they may be involved in accelerating their evolution. This provides one of the very first reports of early steps in neo-sex chromosome differentiation in mammals, and for the first time a cellular framework for the addition-attrition model of sex chromosome evolution.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals.Genes (Basel). 2021 Sep 17;12(9):1434. doi: 10.3390/genes12091434. Genes (Basel). 2021. PMID: 34573416 Free PMC article.
-
Sex chromosome quadrivalents in oocytes of the African pygmy mouse Mus minutoides that harbors non-conventional sex chromosomes.Chromosoma. 2019 Sep;128(3):397-411. doi: 10.1007/s00412-019-00699-4. Epub 2019 Mar 27. Chromosoma. 2019. PMID: 30919035
-
Relationship Between Sequence Homology, Genome Architecture, and Meiotic Behavior of the Sex Chromosomes in North American Voles.Genetics. 2018 Sep;210(1):83-97. doi: 10.1534/genetics.118.301182. Epub 2018 Jul 12. Genetics. 2018. PMID: 30002081 Free PMC article.
-
The origin and evolution of the pseudoautosomal regions of human sex chromosomes.Hum Mol Genet. 1998 Dec;7(13):1991-6. doi: 10.1093/hmg/7.13.1991. Hum Mol Genet. 1998. PMID: 9817914 Review.
-
The evolution of a neo-XY1Y2 sex chromosome system by autosome-sex chromosome fusion in Dundocoris nodulicarinus Jacobs (Heteroptera: Aradidae: Carventinae).Chromosome Res. 2004;12(2):175-91. doi: 10.1023/b:chro.0000013155.99614.57. Chromosome Res. 2004. PMID: 15053487 Review.
Cited by
-
A brief review of vertebrate sex evolution with a pledge for integrative research: towards 'sexomics'.Philos Trans R Soc Lond B Biol Sci. 2021 Aug 30;376(1832):20200426. doi: 10.1098/rstb.2020.0426. Epub 2021 Jul 12. Philos Trans R Soc Lond B Biol Sci. 2021. PMID: 34247497 Free PMC article. Review.
-
Sex differences in the meiotic behavior of an XX sex chromosome pair in males and females of the mole vole Ellobius tancrei: turning an X into a Y chromosome?Chromosoma. 2021 Sep;130(2-3):113-131. doi: 10.1007/s00412-021-00755-y. Epub 2021 Apr 6. Chromosoma. 2021. PMID: 33825031
-
Expanding the classical paradigm: what we have learnt from vertebrates about sex chromosome evolution.Philos Trans R Soc Lond B Biol Sci. 2021 Sep 13;376(1833):20200097. doi: 10.1098/rstb.2020.0097. Epub 2021 Jul 26. Philos Trans R Soc Lond B Biol Sci. 2021. PMID: 34304593 Free PMC article. Review.
-
Strategies for meiotic sex chromosome dynamics and telomeric elongation in Marsupials.PLoS Genet. 2022 Feb 7;18(2):e1010040. doi: 10.1371/journal.pgen.1010040. eCollection 2022 Feb. PLoS Genet. 2022. PMID: 35130272 Free PMC article.
-
Meiotic Behavior of Achiasmate Sex Chromosomes in the African Pygmy Mouse Mus mattheyi Offers New Insights into the Evolution of Sex Chromosome Pairing and Segregation in Mammals.Genes (Basel). 2021 Sep 17;12(9):1434. doi: 10.3390/genes12091434. Genes (Basel). 2021. PMID: 34573416 Free PMC article.
References
-
- Ohno S. Sex chromosomes and sex linked genes. Berlin: Springer; 1967.
-
- Rice WR. Evolution of the Y Sex Chromosome in Animals: Y chromosomes evolve through the degeneration of autosomes. BioScience. 1996;46(5):331–43. 10.2307/1312947 - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
