

NEDD8 and ubiquitin ligation by cullin-RING E3 ligases
- PMID: 33160249
- PMCID: PMC8096640
- DOI: 10.1016/j.sbi.2020.10.007
NEDD8 and ubiquitin ligation by cullin-RING E3 ligases
Abstract
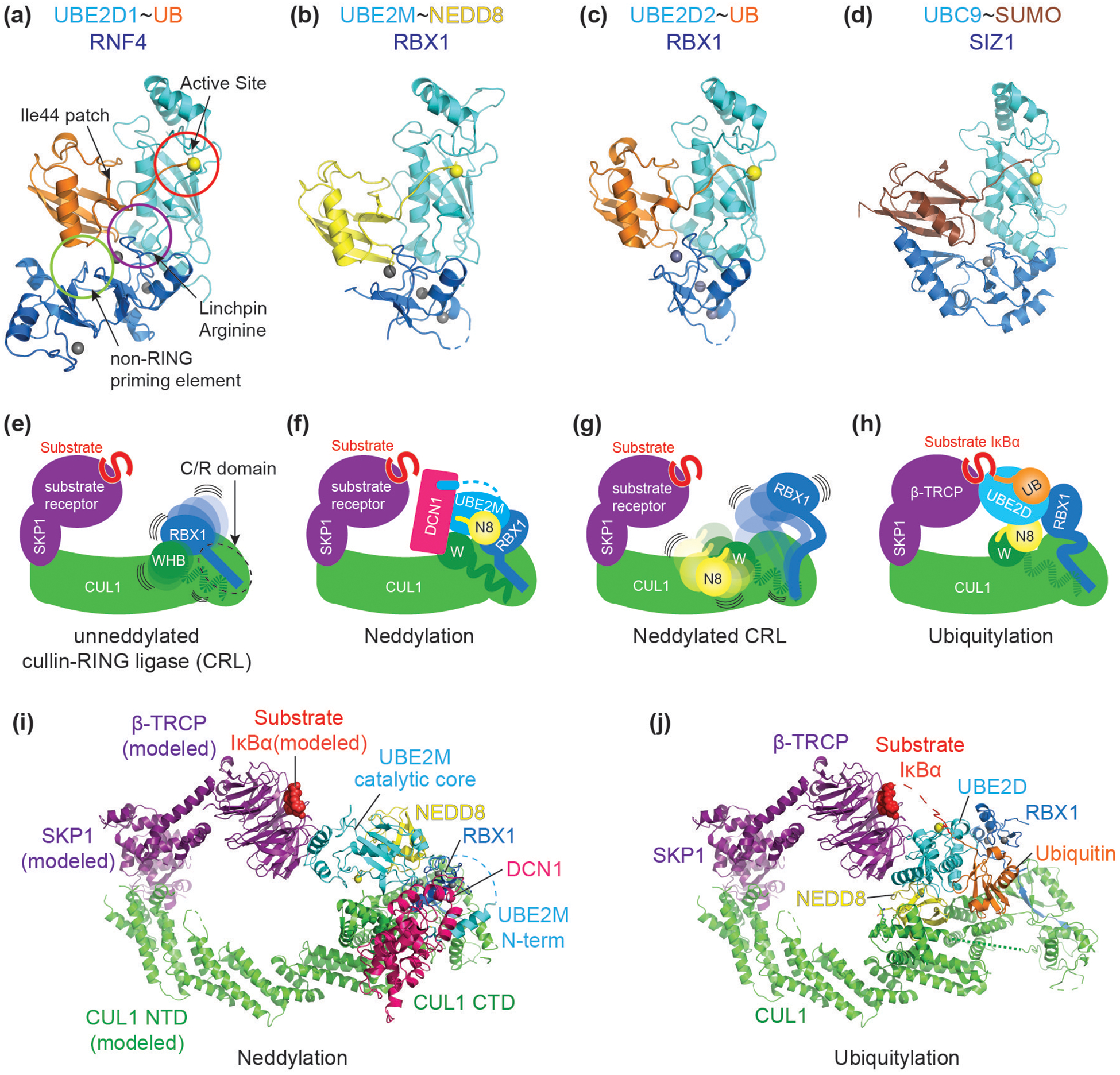
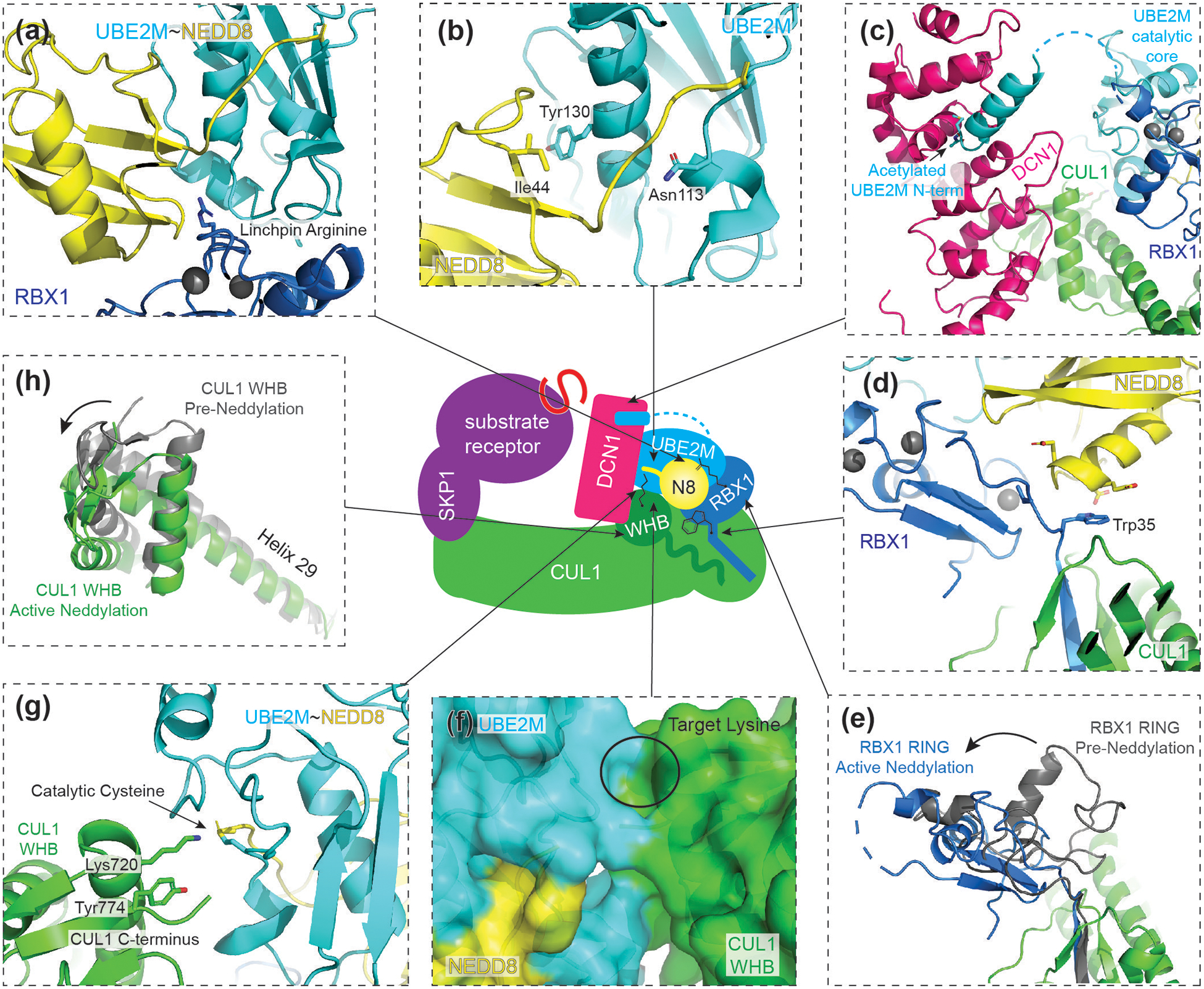
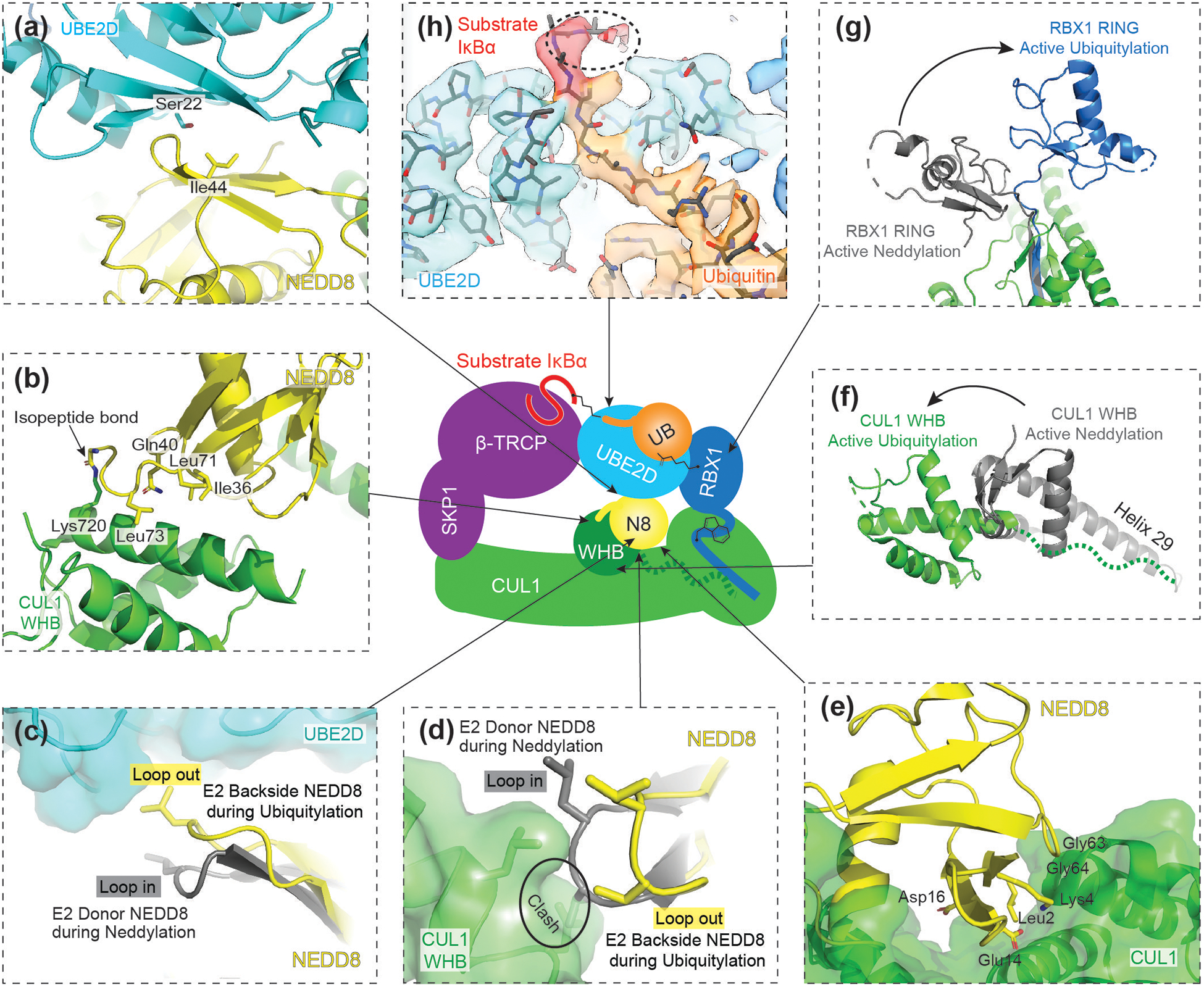
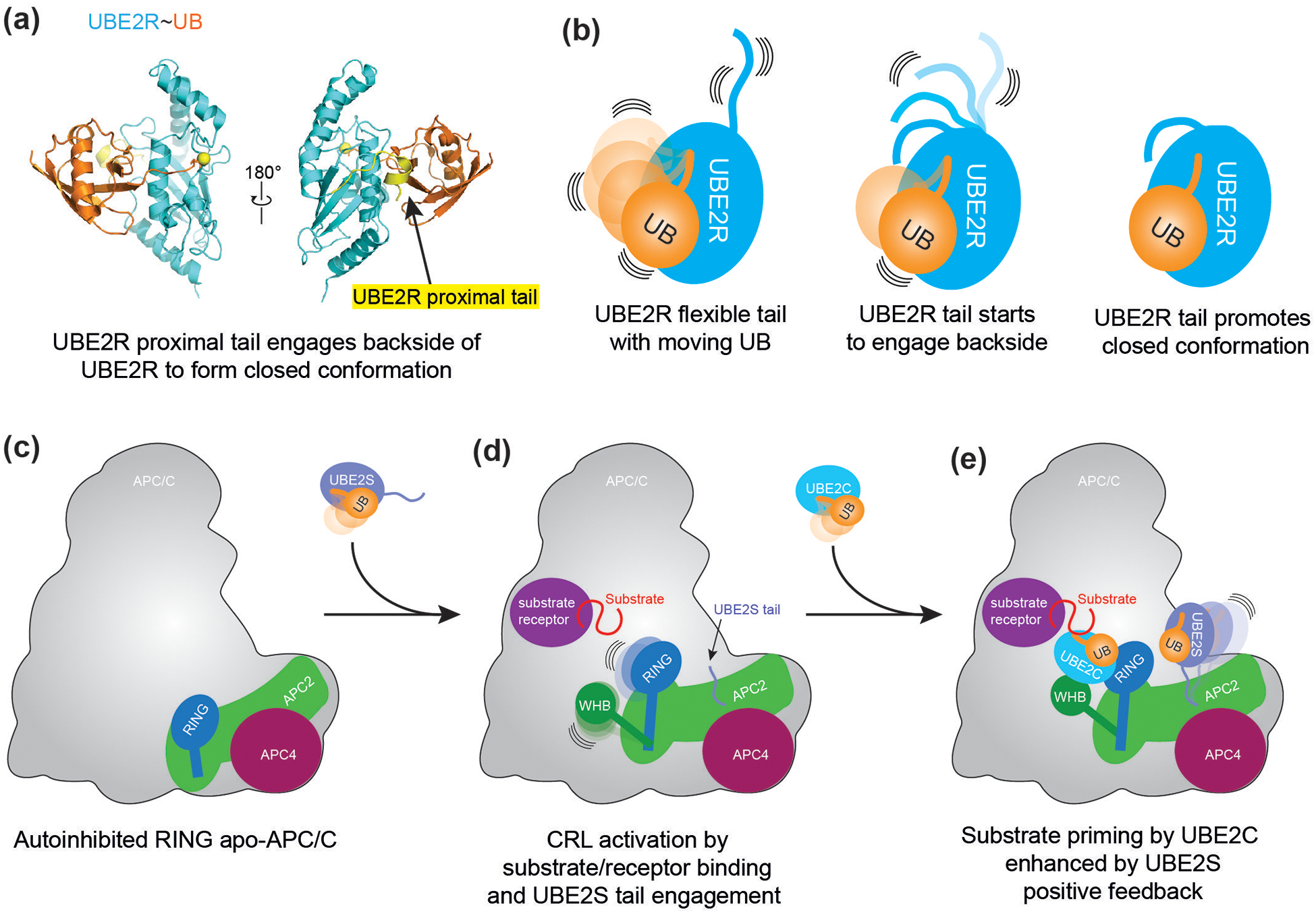
RING E3s comprise the largest family of ubiquitin (UB) and ubiquitin-like protein (UBL) ligases. RING E3s typically promote UB or UBL transfer from the active site of an associated E2 enzyme to a distally-recruited substrate. Many RING E3s - including the cullin-RING ligase family - are multifunctional, interacting with various E2s (or other E3s) to target distinct proteins, transfer different UBLs, or to initially modify substrates with UB or subsequently elongate UB chains. Here we consider recent structures of cullin-RING ligases, and their partner E2 enzymes, representing ligation reactions. The studies collectively reveal multimodal mechanisms - interactions between ancillary E2 or E3 domains, post-translational modifications, or auxiliary binding partners - directing cullin-RING E3-E2 enzyme active sites to modify their specific targets.
Copyright © 2020 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare no competing interest.
Figures
Similar articles
-
Strategies to Trap Enzyme-Substrate Complexes that Mimic Michaelis Intermediates During E3-Mediated Ubiquitin-Like Protein Ligation.Methods Mol Biol. 2018;1844:169-196. doi: 10.1007/978-1-4939-8706-1_12. Methods Mol Biol. 2018. PMID: 30242710 Free PMC article.
-
Two functionally distinct E2/E3 pairs coordinate sequential ubiquitination of a common substrate in Caenorhabditis elegans development.Proc Natl Acad Sci U S A. 2017 Aug 8;114(32):E6576-E6584. doi: 10.1073/pnas.1705060114. Epub 2017 Jul 24. Proc Natl Acad Sci U S A. 2017. PMID: 28739890 Free PMC article.
-
Two Distinct Types of E3 Ligases Work in Unison to Regulate Substrate Ubiquitylation.Cell. 2016 Aug 25;166(5):1198-1214.e24. doi: 10.1016/j.cell.2016.07.027. Cell. 2016. PMID: 27565346 Free PMC article.
-
Regulating the Regulators: Recent Revelations in the Control of E3 Ubiquitin Ligases.J Biol Chem. 2015 Aug 28;290(35):21244-51. doi: 10.1074/jbc.R115.675165. Epub 2015 Jul 17. J Biol Chem. 2015. PMID: 26187467 Free PMC article. Review.
-
Cullin 5-RING E3 ubiquitin ligases, new therapeutic targets?Biochimie. 2016 Mar;122:339-47. doi: 10.1016/j.biochi.2015.08.003. Epub 2015 Aug 4. Biochimie. 2016. PMID: 26253693 Review.
Cited by
-
A non-canonical scaffold-type E3 ligase complex mediates protein UFMylation.EMBO J. 2022 Nov 2;41(21):e111015. doi: 10.15252/embj.2022111015. Epub 2022 Sep 19. EMBO J. 2022. PMID: 36121123 Free PMC article.
-
Sequence and structural variations determining the recruitment of WNK kinases to the KLHL3 E3 ligase.Biochem J. 2022 Mar 18;479(5):661-675. doi: 10.1042/BCJ20220019. Biochem J. 2022. PMID: 35179207 Free PMC article.
-
Functional conservation and divergence of the helix-turn-helix motif of E2 ubiquitin-conjugating enzymes.EMBO J. 2022 Feb 1;41(3):e108823. doi: 10.15252/embj.2021108823. Epub 2021 Dec 23. EMBO J. 2022. PMID: 34942047 Free PMC article.
-
The C-terminal tail of CSNAP attenuates the CSN complex.Life Sci Alliance. 2023 Jul 17;6(10):e202201634. doi: 10.26508/lsa.202201634. Print 2023 Oct. Life Sci Alliance. 2023. PMID: 37460146 Free PMC article.
-
Insight into Viral Hijacking of CRL4 Ubiquitin Ligase through Structural Analysis of the pUL145-DDB1 Complex.J Virol. 2022 Sep 14;96(17):e0082622. doi: 10.1128/jvi.00826-22. Epub 2022 Aug 8. J Virol. 2022. PMID: 35938868 Free PMC article.
References
-
- Pan ZQ, Kentsis A, Dias DC, Yamoah K, Wu K: Nedd8 on cullin: building an expressway to protein destruction. Oncogene 2004, 23:1985–1997. - PubMed
-
- Komander D, Rape M: The ubiquitin code. Annu Rev Biochem 2012, 81:203–229. - PubMed
-
- Pruneda JN, Littlefield PJ, Soss SE, Nordquist KA, Chazin WJ, Brzovic PS, Klevit RE: Structure of an E3:E2~Ub complex reveals an allosteric mechanism shared among RING/U-box ligases. Mol Cell 2012, 47:933–942. - PMC - PubMed
-
A NMR-based study and structural model defining a key hydrogen bond between a conserved side chain within E3s and carbonyl of E2s for stabilizing the closed E2~UB conformation.
-
- Hamilton KS, Ellison MJ, Barber KR, Williams RS, Huzil JT, McKenna S, Ptak C, Glover M, Shaw GS: Structure of a conjugating enzyme-ubiquitin thiolester intermediate reveals a novel role for the ubiquitin tail. Structure 2001, 9:897–904. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
