

Endoskeletal mineralization in chimaera and a comparative guide to tessellated cartilage in chondrichthyan fishes (sharks, rays and chimaera)
- PMID: 33050779
- PMCID: PMC7653374
- DOI: 10.1098/rsif.2020.0474
Endoskeletal mineralization in chimaera and a comparative guide to tessellated cartilage in chondrichthyan fishes (sharks, rays and chimaera)
Abstract
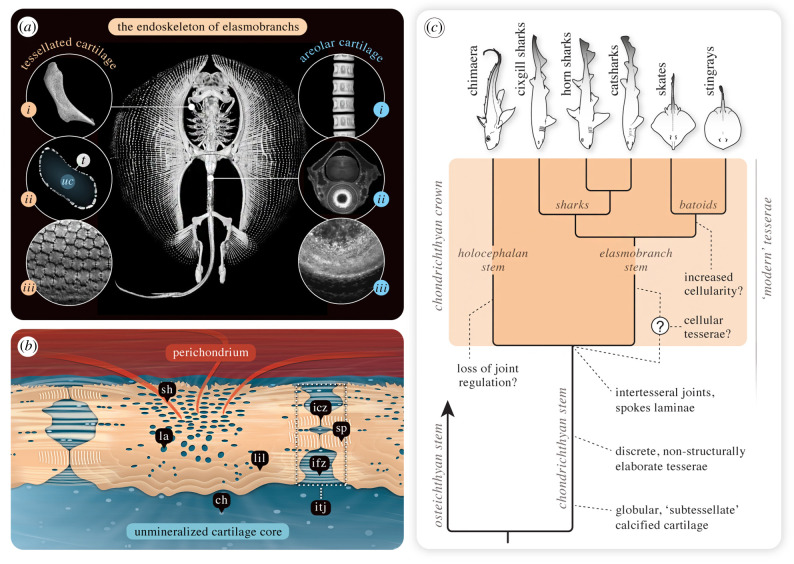
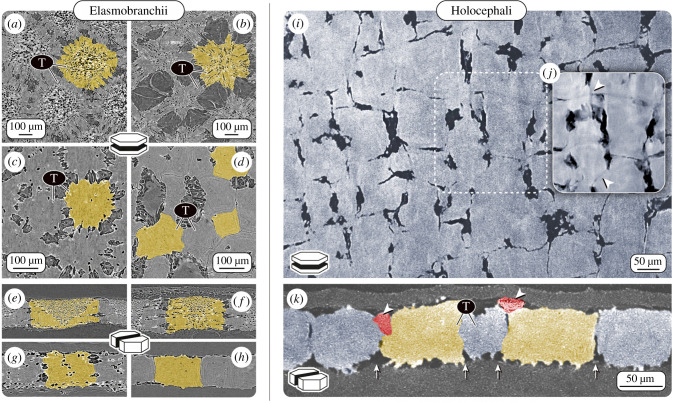
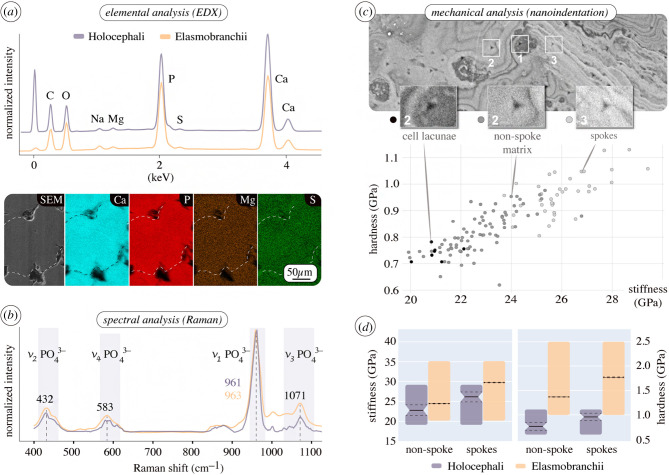
An accepted uniting character of modern cartilaginous fishes (sharks, rays, chimaera) is the presence of a mineralized, skeletal crust, tiled by numerous minute plates called tesserae. Tesserae have, however, never been demonstrated in modern chimaera and it is debated whether the skeleton mineralizes at all. We show for the first time that tessellated cartilage was not lost in chimaera, as has been previously postulated, and is in many ways similar to that of sharks and rays. Tesserae in Chimaera monstrosa are less regular in shape and size in comparison to the general scheme of polygonal tesserae in sharks and rays, yet share several features with them. For example, Chimaera tesserae, like those of elasmobranchs, possess both intertesseral joints (unmineralized regions, where fibrous tissue links adjacent tesserae) and recurring patterns of local mineral density variation (e.g. Liesegang lines, hypermineralized 'spokes'), reflecting periodic accretion of mineral at tesseral edges as tesserae grow. Chimaera monstrosa's tesserae, however, appear to lack the internal cell networks that characterize tesserae in elasmobranchs, indicating fundamental differences among chondrichthyan groups in how calcification is controlled. By compiling and comparing recent ultrastructure data on tesserae, we also provide a synthesized, up-to-date and comparative glossary on tessellated cartilage, as well as a perspective on the current state of research into the topic, offering benchmark context for future research into modern and extinct vertebrate skeletal tissues.
Keywords: biomineralization; cartilaginous fish; tessellated cartilage; tesserae; vertebrate endoskeleton.
Conflict of interest statement
We declare we have no competing interests.
Figures


Similar articles
-
Ultrastructural and developmental features of the tessellated endoskeleton of elasmobranchs (sharks and rays).J Anat. 2016 Nov;229(5):681-702. doi: 10.1111/joa.12508. Epub 2016 Aug 24. J Anat. 2016. PMID: 27557870 Free PMC article.
-
Calcified cartilage or bone? Collagens in the tessellated endoskeletons of cartilaginous fish (sharks and rays).J Struct Biol. 2017 Oct;200(1):54-71. doi: 10.1016/j.jsb.2017.09.005. Epub 2017 Sep 18. J Struct Biol. 2017. PMID: 28923317
-
Mechanical properties of stingray tesserae: High-resolution correlative analysis of mineral density and indentation moduli in tessellated cartilage.Acta Biomater. 2019 Sep 15;96:421-435. doi: 10.1016/j.actbio.2019.06.038. Epub 2019 Jun 27. Acta Biomater. 2019. PMID: 31254686
-
Architectural and ultrastructural features of tessellated calcified cartilage in modern and extinct chondrichthyan fishes.J Fish Biol. 2021 Apr;98(4):919-941. doi: 10.1111/jfb.14376. Epub 2020 Jul 1. J Fish Biol. 2021. PMID: 32388865 Review.
-
Mineral homeostasis and regulation of mineralization processes in the skeletons of sharks, rays and relatives (Elasmobranchii).Semin Cell Dev Biol. 2015 Oct;46:51-67. doi: 10.1016/j.semcdb.2015.10.022. Epub 2015 Nov 10. Semin Cell Dev Biol. 2015. PMID: 26546857 Review.
Cited by
-
Morphology of joints and patterns of cartilage calcification in the endoskeleton of the batoid Raja cf. polystigma.J Anat. 2022 Jun;240(6):1127-1140. doi: 10.1111/joa.13623. Epub 2022 Jan 17. J Anat. 2022. PMID: 35037257 Free PMC article.
-
A new Meckel's cartilage from the Devonian Hangenberg black shale in Morocco and its position in chondrichthyan jaw morphospace.PeerJ. 2022 Dec 21;10:e14418. doi: 10.7717/peerj.14418. eCollection 2022. PeerJ. 2022. PMID: 36573235 Free PMC article.
-
Mineralization of the Callorhinchus Vertebral Column (Holocephali; Chondrichthyes).Front Genet. 2020 Nov 26;11:571694. doi: 10.3389/fgene.2020.571694. eCollection 2020. Front Genet. 2020. PMID: 33329708 Free PMC article.
-
Mineralized Cartilage and Bone-Like Tissues in Chondrichthyans Offer Potential Insights Into the Evolution and Development of Mineralized Tissues in the Vertebrate Endoskeleton.Front Genet. 2021 Dec 22;12:762042. doi: 10.3389/fgene.2021.762042. eCollection 2021. Front Genet. 2021. PMID: 35003210 Free PMC article. Review.
-
The combined cartilage growth - calcification patterns in the wing-fins of Rajidae (Chondrichthyes): A divergent model from endochondral ossification of tetrapods.Microsc Res Tech. 2022 Nov;85(11):3642-3652. doi: 10.1002/jemt.24217. Epub 2022 Aug 3. Microsc Res Tech. 2022. PMID: 36250446 Free PMC article.
References
-
- Hall BK. 2005. Bones and cartilage: developmental skeletal biology. London, UK: Academic Press.
-
- Applegate SP. 1967. A survey of shark hard parts, pp. 37–66. Baltimore, MD: Johns Hopkins University Press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
