

SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies
- PMID: 33045718
- PMCID: PMC8092461
- DOI: 10.1038/s41586-020-2852-1
SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies
Erratum in
-
Author Correction: SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies.Nature. 2024 Apr;628(8008):E2. doi: 10.1038/s41586-024-07344-w. Nature. 2024. PMID: 38565636 No abstract available.
Abstract
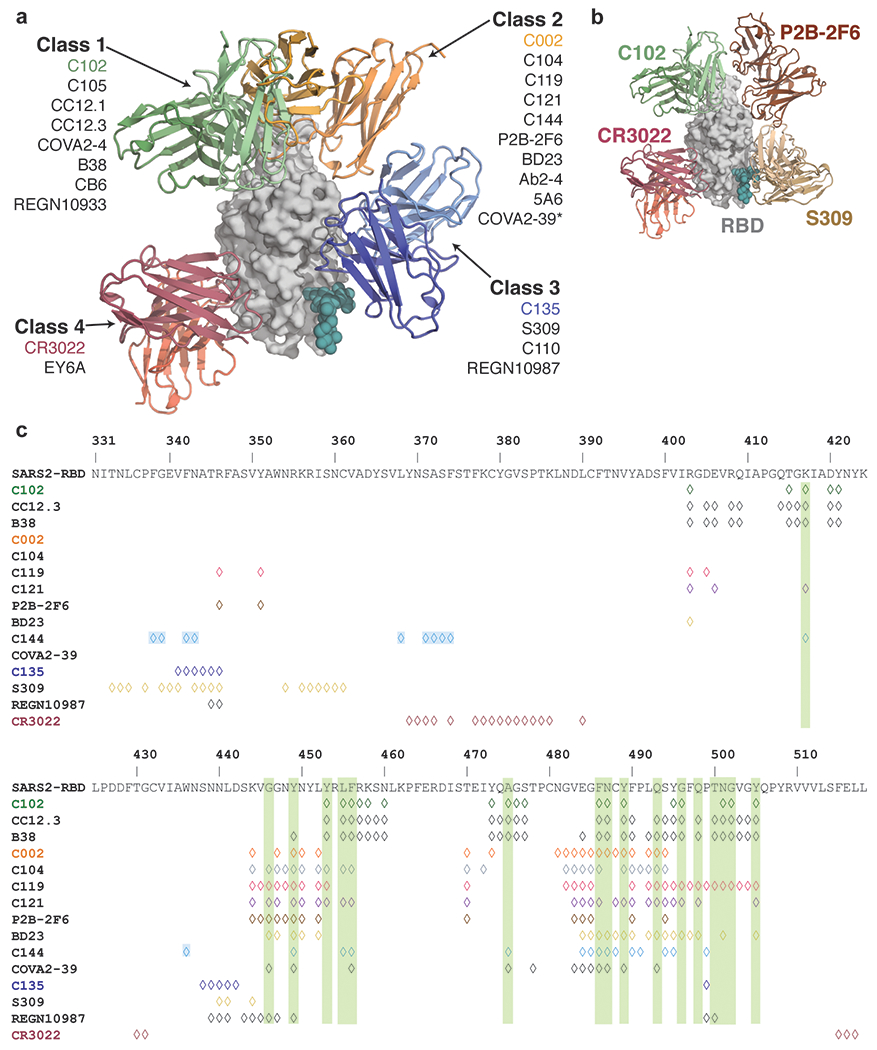
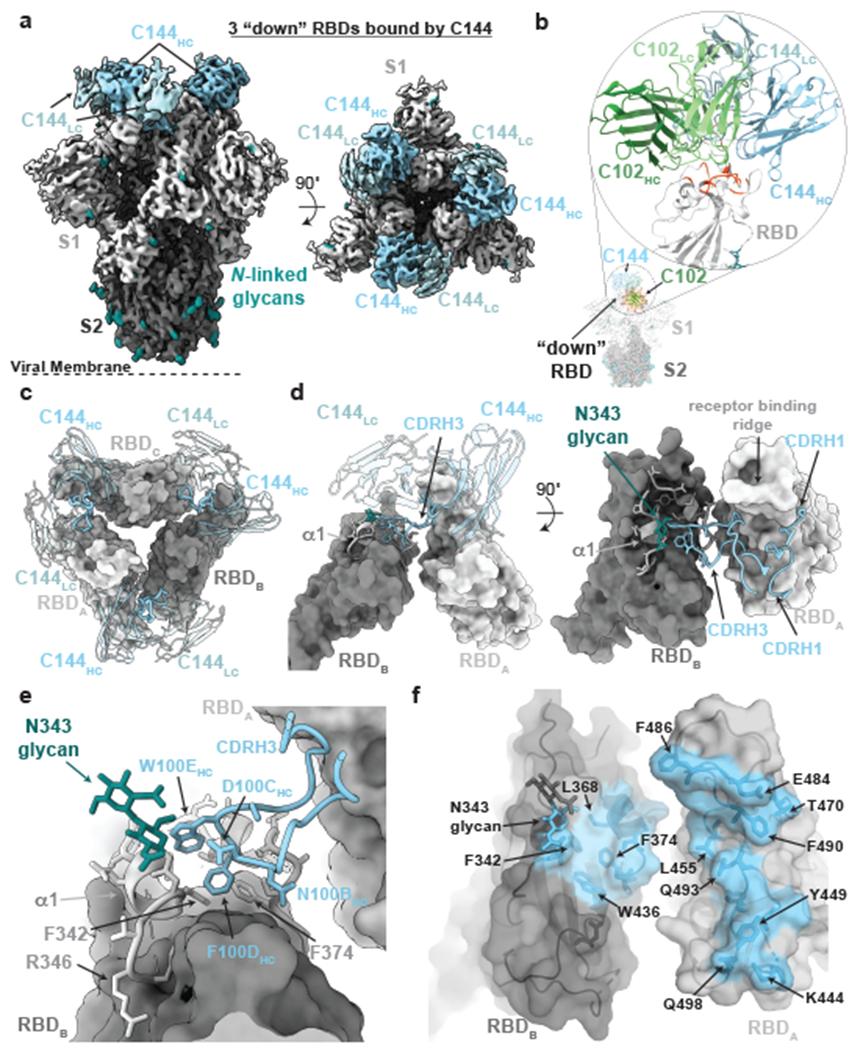
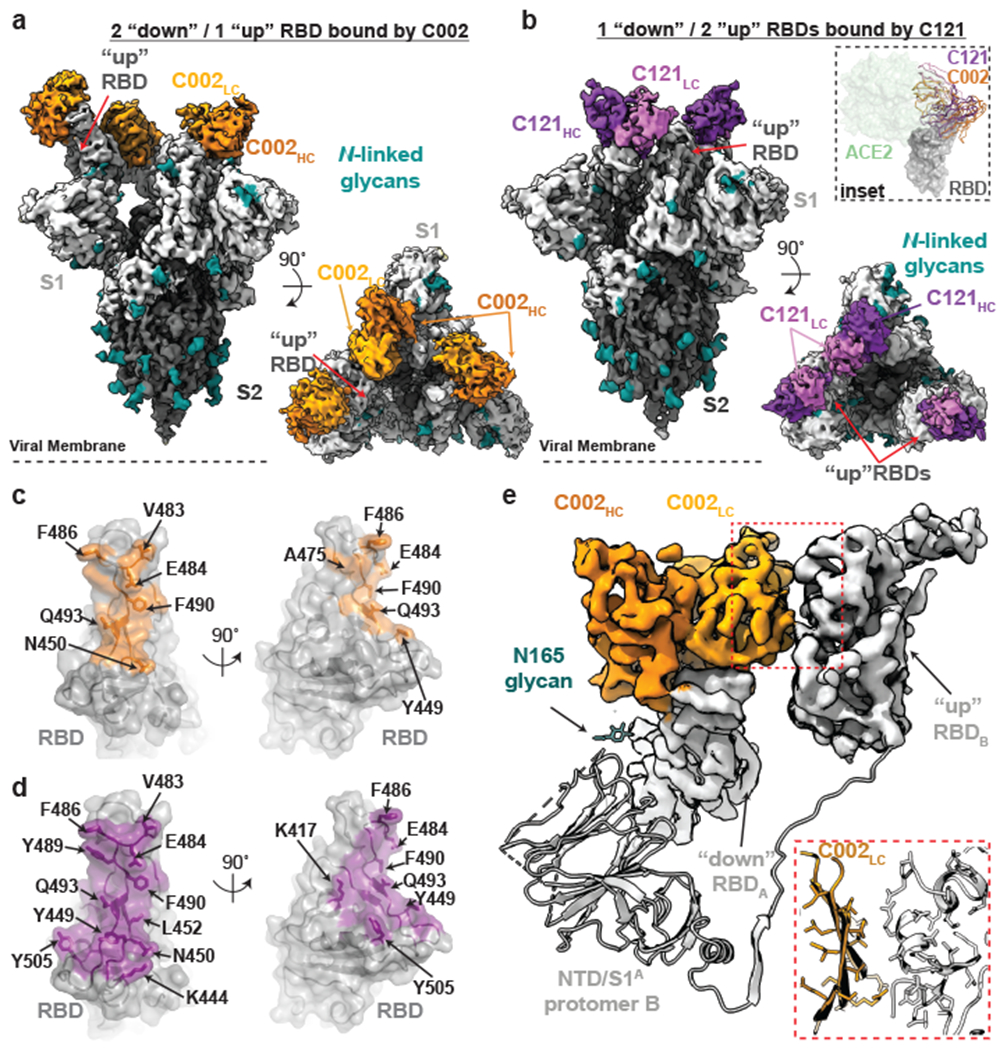
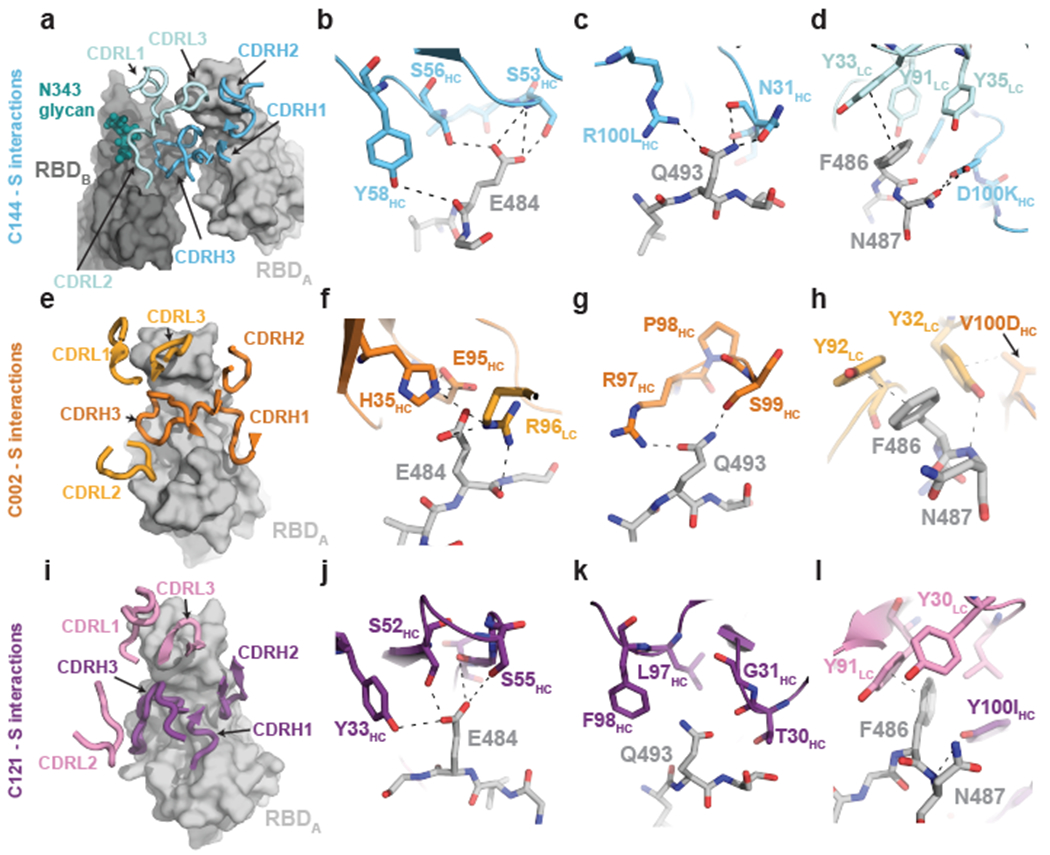
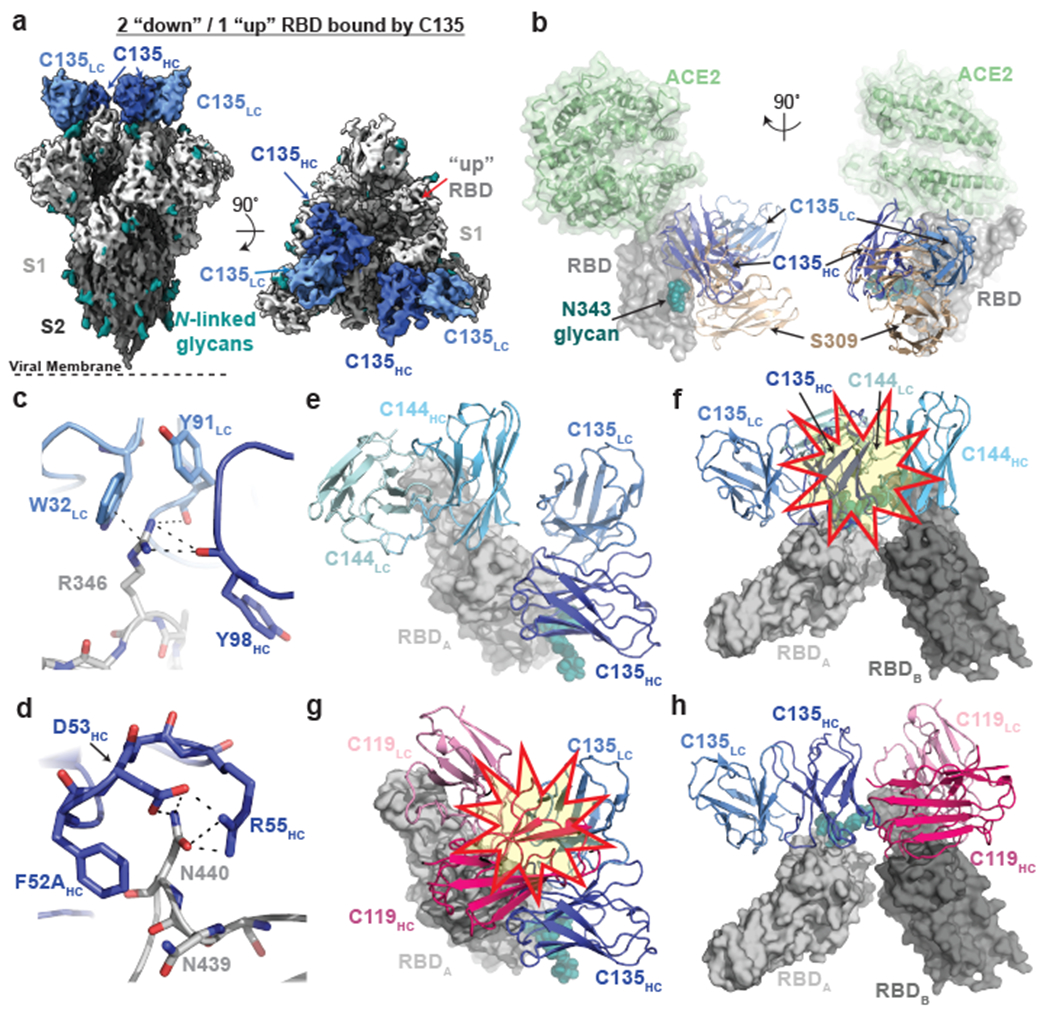
The coronavirus disease 2019 (COVID-19) pandemic presents an urgent health crisis. Human neutralizing antibodies that target the host ACE2 receptor-binding domain (RBD) of the severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) spike protein1-5 show promise therapeutically and are being evaluated clinically6-8. Here, to identify the structural correlates of SARS-CoV-2 neutralization, we solved eight new structures of distinct COVID-19 human neutralizing antibodies5 in complex with the SARS-CoV-2 spike trimer or RBD. Structural comparisons allowed us to classify the antibodies into categories: (1) neutralizing antibodies encoded by the VH3-53 gene segment with short CDRH3 loops that block ACE2 and bind only to 'up' RBDs; (2) ACE2-blocking neutralizing antibodies that bind both up and 'down' RBDs and can contact adjacent RBDs; (3) neutralizing antibodies that bind outside the ACE2 site and recognize both up and down RBDs; and (4) previously described antibodies that do not block ACE2 and bind only to up RBDs9. Class 2 contained four neutralizing antibodies with epitopes that bridged RBDs, including a VH3-53 antibody that used a long CDRH3 with a hydrophobic tip to bridge between adjacent down RBDs, thereby locking the spike into a closed conformation. Epitope and paratope mapping revealed few interactions with host-derived N-glycans and minor contributions of antibody somatic hypermutations to epitope contacts. Affinity measurements and mapping of naturally occurring and in vitro-selected spike mutants in 3D provided insight into the potential for SARS-CoV-2 to escape from antibodies elicited during infection or delivered therapeutically. These classifications and structural analyses provide rules for assigning current and future human RBD-targeting antibodies into classes, evaluating avidity effects and suggesting combinations for clinical use, and provide insight into immune responses against SARS-CoV-2.
Conflict of interest statement
Competing interests
The Rockefeller University has filed a provisional patent application in connection with this work on which D.F.R. and M.C.N. are inventors (US 63/021,387).
Figures
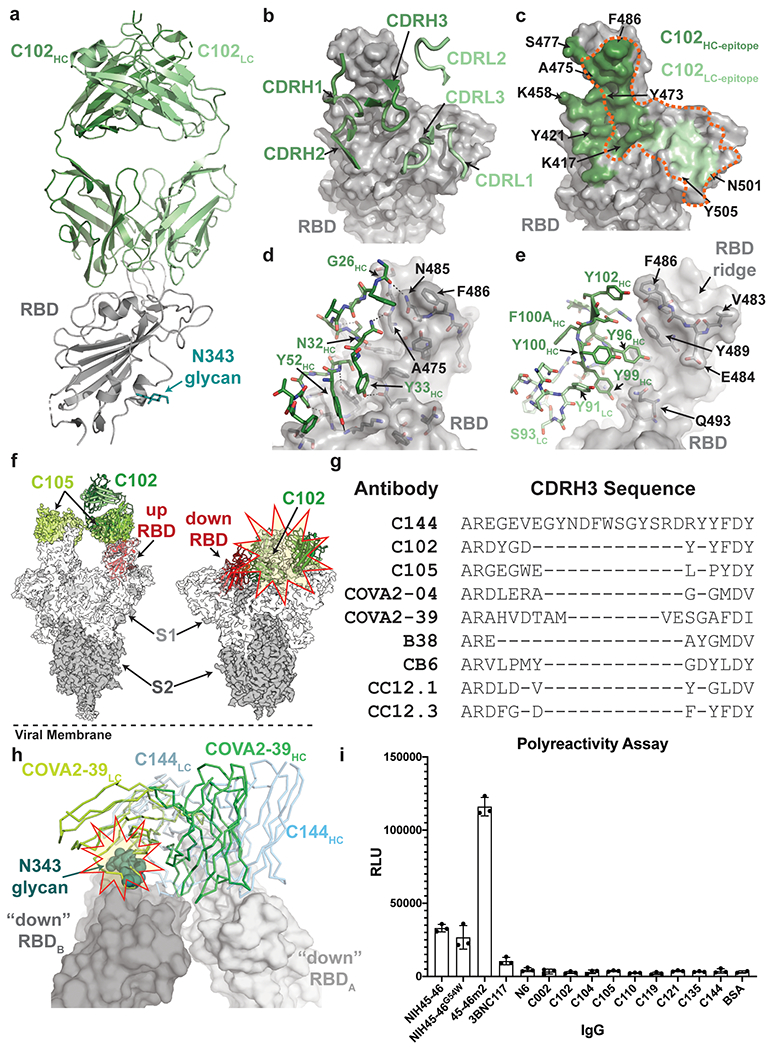
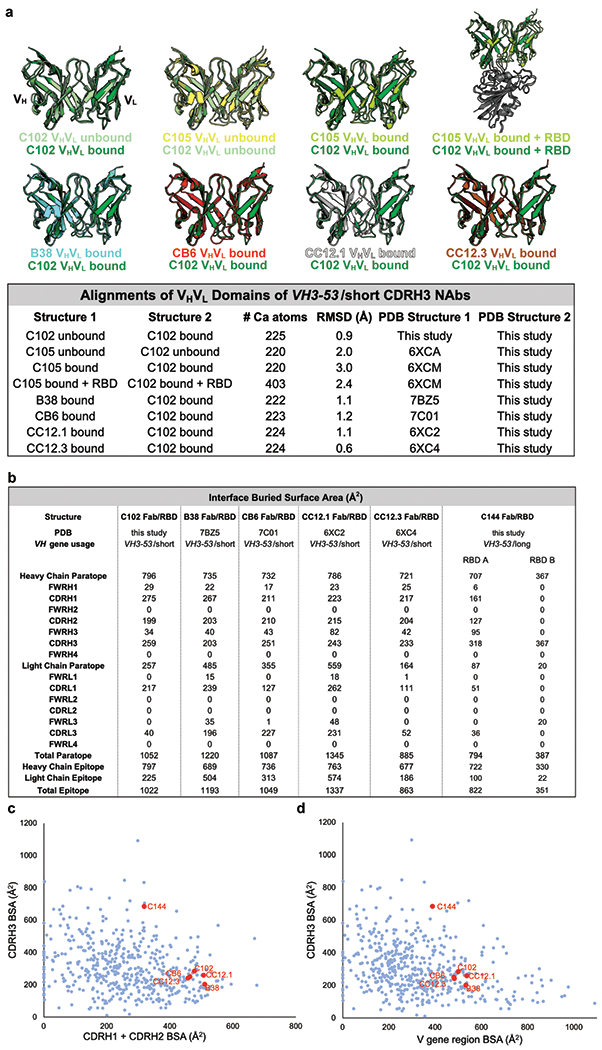

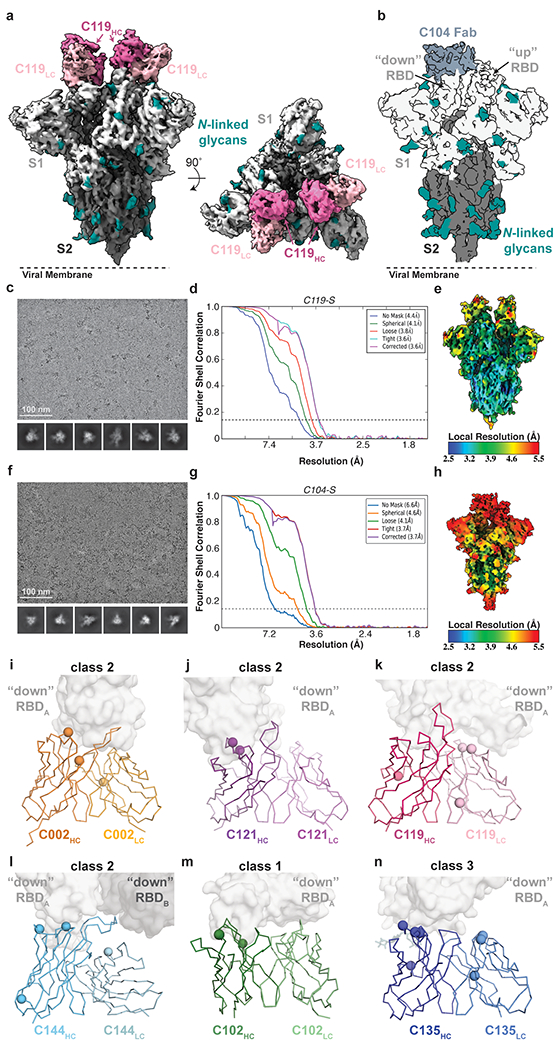
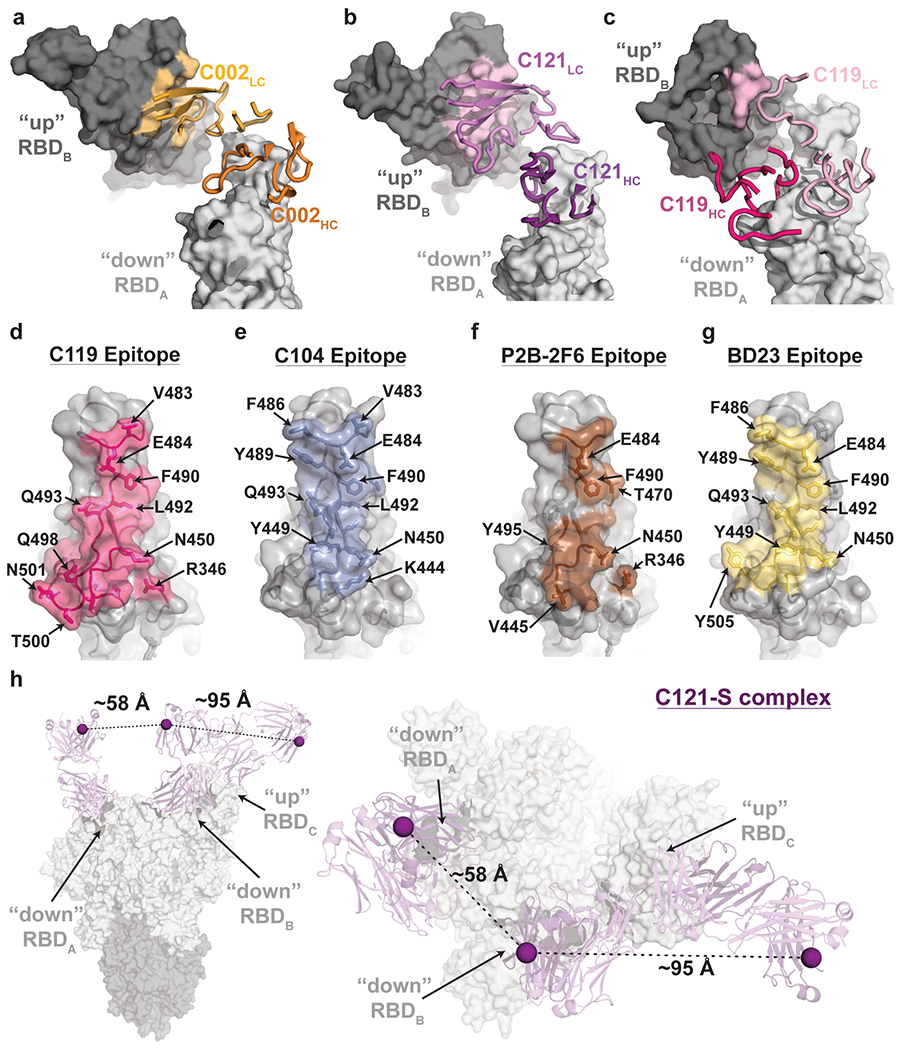



Update of
-
Structural classification of neutralizing antibodies against the SARS-CoV-2 spike receptor-binding domain suggests vaccine and therapeutic strategies.bioRxiv [Preprint]. 2020 Aug 30:2020.08.30.273920. doi: 10.1101/2020.08.30.273920. bioRxiv. 2020. Update in: Nature. 2020 Dec;588(7839):682-687. doi: 10.1038/s41586-020-2852-1. PMID: 32869026 Free PMC article. Updated. Preprint.
Similar articles
-
Structural classification of neutralizing antibodies against the SARS-CoV-2 spike receptor-binding domain suggests vaccine and therapeutic strategies.bioRxiv [Preprint]. 2020 Aug 30:2020.08.30.273920. doi: 10.1101/2020.08.30.273920. bioRxiv. 2020. Update in: Nature. 2020 Dec;588(7839):682-687. doi: 10.1038/s41586-020-2852-1. PMID: 32869026 Free PMC article. Updated. Preprint.
-
Structural basis and analysis of hamster ACE2 binding to different SARS-CoV-2 spike RBDs.J Virol. 2024 Mar 19;98(3):e0115723. doi: 10.1128/jvi.01157-23. Epub 2024 Feb 2. J Virol. 2024. PMID: 38305152 Free PMC article.
-
Broad cross-reactivity across sarbecoviruses exhibited by a subset of COVID-19 donor-derived neutralizing antibodies.Cell Rep. 2021 Sep 28;36(13):109760. doi: 10.1016/j.celrep.2021.109760. Epub 2021 Sep 8. Cell Rep. 2021. PMID: 34534459 Free PMC article.
-
Structural basis of severe acute respiratory syndrome coronavirus 2 infection.Curr Opin HIV AIDS. 2021 Jan;16(1):74-81. doi: 10.1097/COH.0000000000000658. Curr Opin HIV AIDS. 2021. PMID: 33186231 Review.
-
Distinct Conformations of SARS-CoV-2 Omicron Spike Protein and Its Interaction with ACE2 and Antibody.Int J Mol Sci. 2023 Feb 14;24(4):3774. doi: 10.3390/ijms24043774. Int J Mol Sci. 2023. PMID: 36835186 Free PMC article. Review.
Cited by
-
Engineering a SARS-CoV-2 Vaccine Targeting the Receptor-Binding Domain Cryptic-Face via Immunofocusing.ACS Cent Sci. 2024 Sep 17;10(10):1871-1884. doi: 10.1021/acscentsci.4c00722. eCollection 2024 Oct 23. ACS Cent Sci. 2024. PMID: 39463836 Free PMC article.
-
Monoclonal antibodies: From magic bullet to precision weapon.Mol Biomed. 2024 Oct 11;5(1):47. doi: 10.1186/s43556-024-00210-1. Mol Biomed. 2024. PMID: 39390211 Free PMC article. Review.
-
Comparison of the Neutralization Power of Sotrovimab Against SARS-CoV-2 Variants: Development of a Rapid Computational Method.JMIR Bioinform Biotechnol. 2024 Oct 10;5:e58018. doi: 10.2196/58018. JMIR Bioinform Biotechnol. 2024. PMID: 39388246 Free PMC article.
-
Structural basis for receptor-binding domain mobility of the spike in SARS-CoV-2 BA.2.86 and JN.1.Nat Commun. 2024 Oct 7;15(1):8574. doi: 10.1038/s41467-024-52808-2. Nat Commun. 2024. PMID: 39375326 Free PMC article.
-
Exploring the ability of the MD+FoldX method to predict SARS-CoV-2 antibody escape mutations using large-scale data.Sci Rep. 2024 Oct 4;14(1):23122. doi: 10.1038/s41598-024-72491-z. Sci Rep. 2024. PMID: 39366988 Free PMC article.
References
Additional References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
