

More than just a ticket canceller: the mitochondrial processing peptidase tailors complex precursor proteins at internal cleavage sites
- PMID: 32997570
- PMCID: PMC8734313
- DOI: 10.1091/mbc.E20-08-0524
More than just a ticket canceller: the mitochondrial processing peptidase tailors complex precursor proteins at internal cleavage sites
Abstract
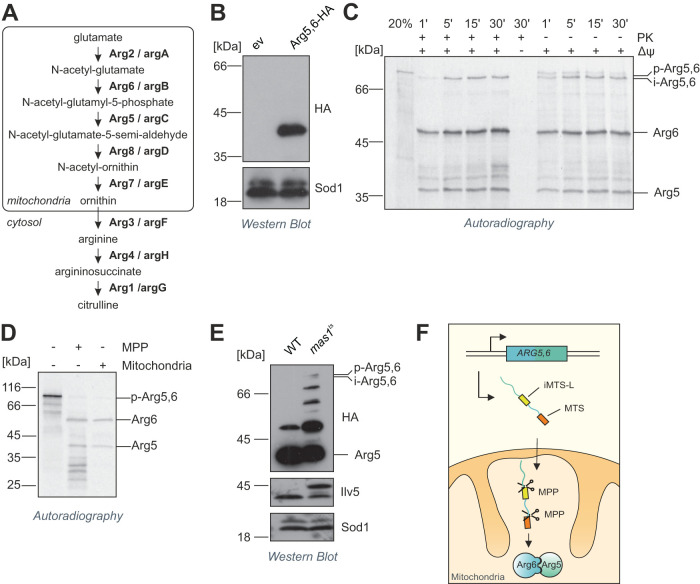
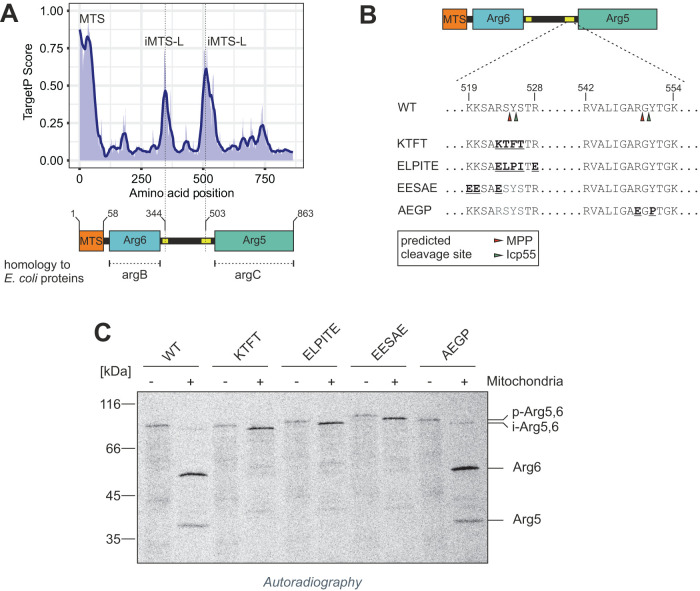
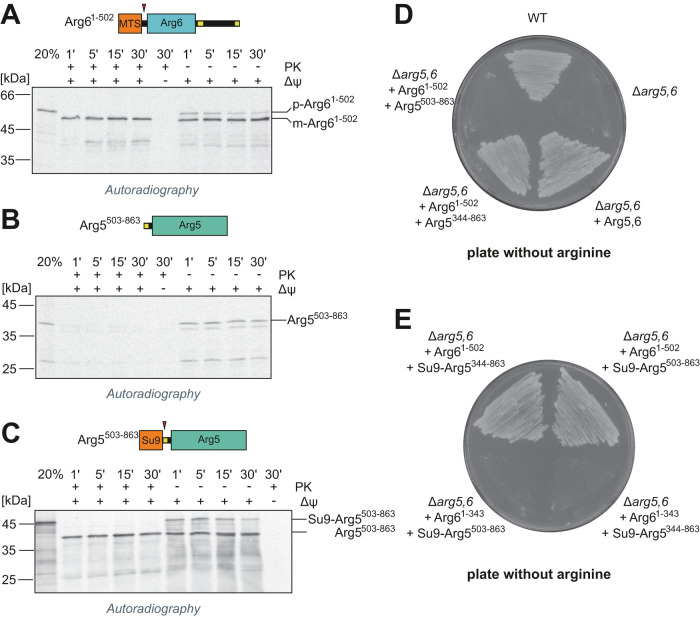
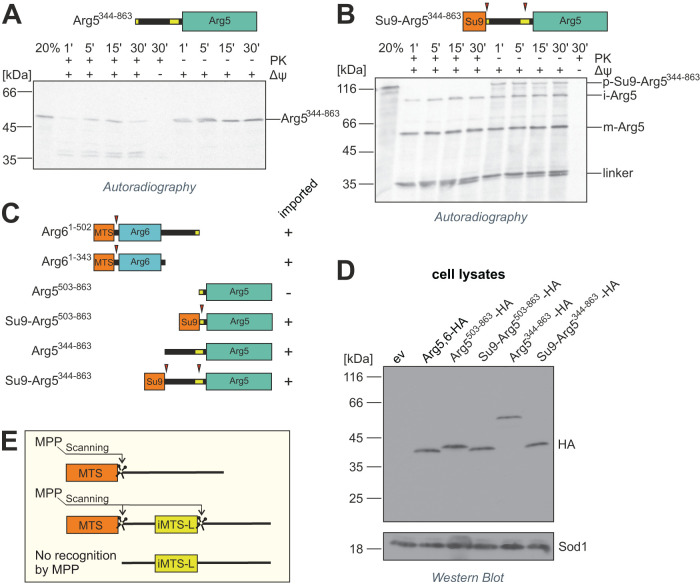
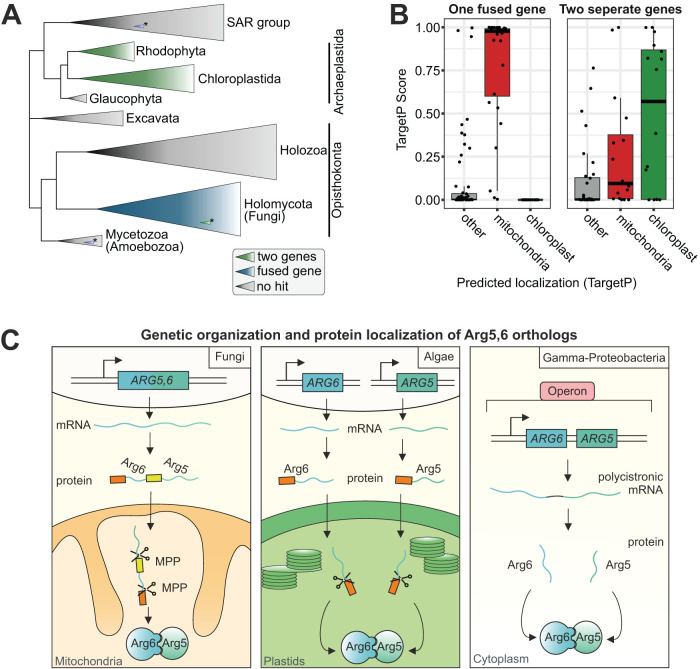
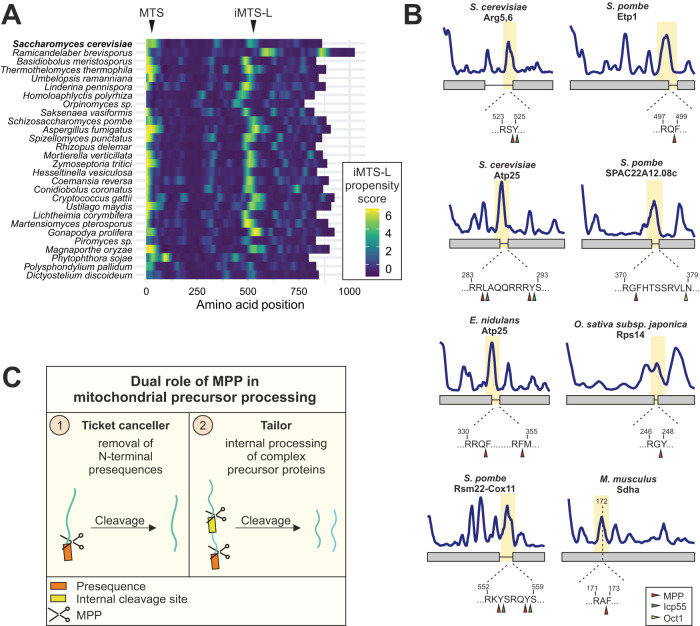
Most mitochondrial proteins are synthesized as precursors that carry N-terminal presequences. After they are imported into mitochondria, these targeting signals are cleaved off by the mitochondrial processing peptidase (MPP). Using the mitochondrial tandem protein Arg5,6 as a model substrate, we demonstrate that MPP has an additional role in preprotein maturation, beyond the removal of presequences. Arg5,6 is synthesized as a polyprotein precursor that is imported into mitochondria and subsequently separated into two distinct enzymes. This internal processing is performed by MPP, which cleaves the Arg5,6 precursor at its N-terminus and at an internal site. The peculiar organization of Arg5,6 is conserved across fungi and reflects the polycistronic arginine operon in prokaryotes. MPP cleavage sites are also present in other mitochondrial fusion proteins from fungi, plants, and animals. Hence, besides its role as a "ticket canceller" for removal of presequences, MPP exhibits a second conserved activity as an internal processing peptidase for complex mitochondrial precursor proteins.
Figures
Similar articles
-
Mitochondrial protein turnover: role of the precursor intermediate peptidase Oct1 in protein stabilization.Mol Biol Cell. 2011 Jul 1;22(13):2135-43. doi: 10.1091/mbc.E11-02-0169. Epub 2011 Apr 27. Mol Biol Cell. 2011. PMID: 21525245 Free PMC article.
-
Processing of mitochondrial precursor proteins.Biomed Biochim Acta. 1991;50(4-6):403-12. Biomed Biochim Acta. 1991. PMID: 1839352 Review.
-
Prediction and identification of new natural substrates of the yeast mitochondrial intermediate peptidase.J Biol Chem. 1995 Nov 10;270(45):27366-73. doi: 10.1074/jbc.270.45.27366. J Biol Chem. 1995. PMID: 7593000
-
Quantitative Profiling for Substrates of the Mitochondrial Presequence Processing Protease Reveals a Set of Nonsubstrate Proteins Increased upon Proteotoxic Stress.J Proteome Res. 2015 Nov 6;14(11):4550-63. doi: 10.1021/acs.jproteome.5b00327. Epub 2015 Oct 21. J Proteome Res. 2015. PMID: 26446170
-
Mitochondrial processing peptidases.Biochim Biophys Acta. 2002 Sep 2;1592(1):63-77. doi: 10.1016/s0167-4889(02)00265-3. Biochim Biophys Acta. 2002. PMID: 12191769 Review.
Cited by
-
Protein Processing in Plant Mitochondria Compared to Yeast and Mammals.Front Plant Sci. 2022 Feb 2;13:824080. doi: 10.3389/fpls.2022.824080. eCollection 2022. Front Plant Sci. 2022. PMID: 35185991 Free PMC article. Review.
-
In-Depth Characterization of Apoptosis N-Terminome Reveals a Link Between Caspase-3 Cleavage and Posttranslational N-Terminal Acetylation.Mol Cell Proteomics. 2023 Jul;22(7):100584. doi: 10.1016/j.mcpro.2023.100584. Epub 2023 May 24. Mol Cell Proteomics. 2023. PMID: 37236440 Free PMC article. Review.
-
Cell-Based Sensors for the Detection of EGF and EGF-Stimulated Ca2+ Signaling.Biosensors (Basel). 2023 Mar 14;13(3):383. doi: 10.3390/bios13030383. Biosensors (Basel). 2023. PMID: 36979595 Free PMC article.
-
Mitochondrial Processing Peptidases-Structure, Function and the Role in Human Diseases.Int J Mol Sci. 2022 Jan 24;23(3):1297. doi: 10.3390/ijms23031297. Int J Mol Sci. 2022. PMID: 35163221 Free PMC article. Review.
-
MTSviewer: A database to visualize mitochondrial targeting sequences, cleavage sites, and mutations on protein structures.PLoS One. 2023 Apr 24;18(4):e0284541. doi: 10.1371/journal.pone.0284541. eCollection 2023. PLoS One. 2023. PMID: 37093842 Free PMC article.
References
-
- Abadjieva A, Pauwels K, Hilven P, Crabeel M (2001). A new yeast metabolon involving at least the two first enzymes of arginine biosynthesis: acetylglutamate synthase activity requires complex formation with acetylglutamate kinase. J Biol Chem 276, 42869–42880. - PubMed
-
- Becker T, Song J, Pfanner N (2019). Versatility of preprotein transfer from the cytosol to mitochondria. Trends Cell Biol 29, 534–548. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
