

Functions of Gle1 are governed by two distinct modes of self-association
- PMID: 32981894
- PMCID: PMC7864074
- DOI: 10.1074/jbc.RA120.015715
Functions of Gle1 are governed by two distinct modes of self-association
Abstract
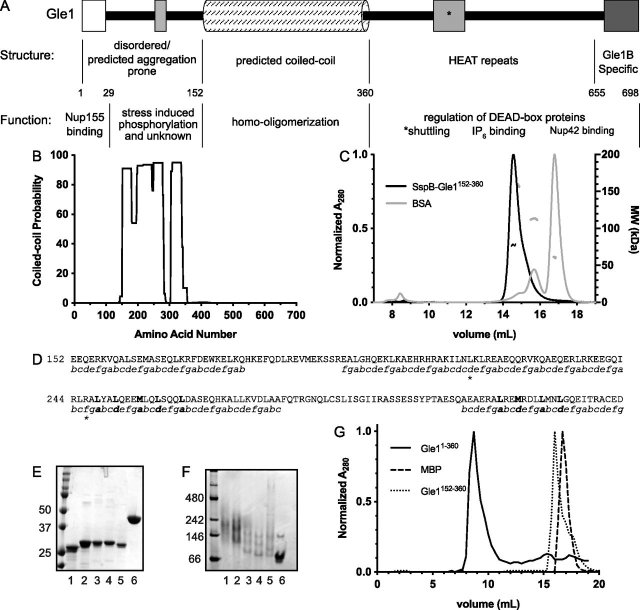
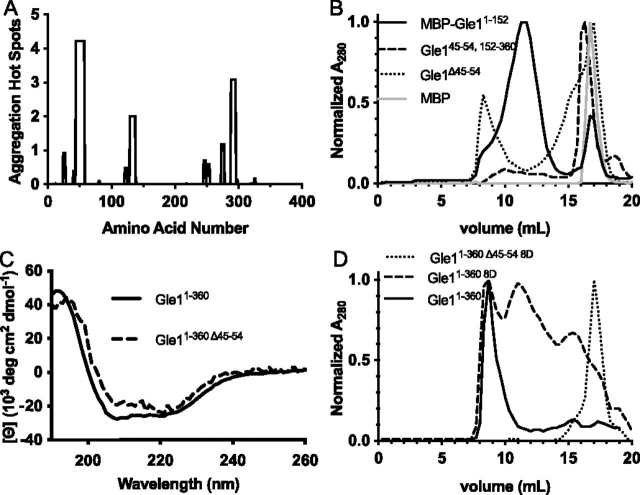
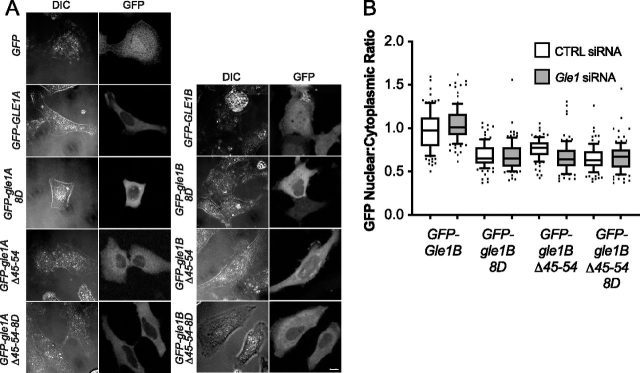
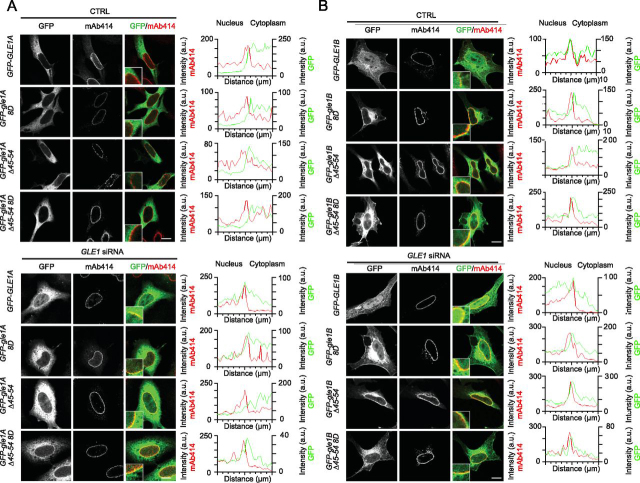
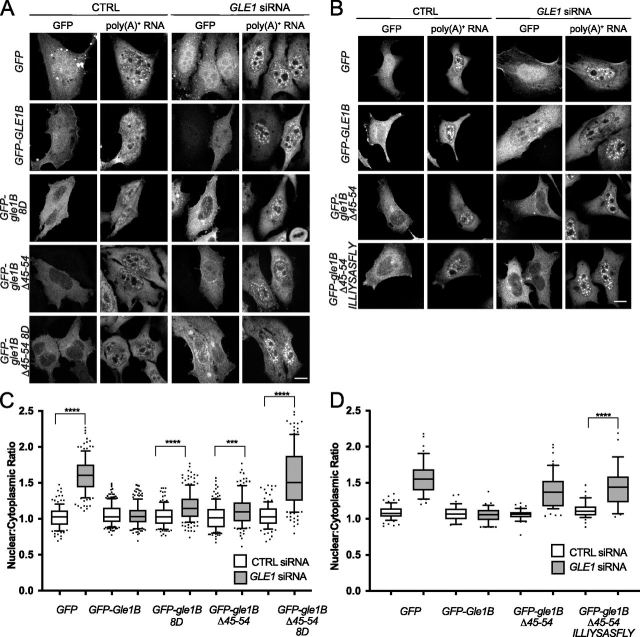
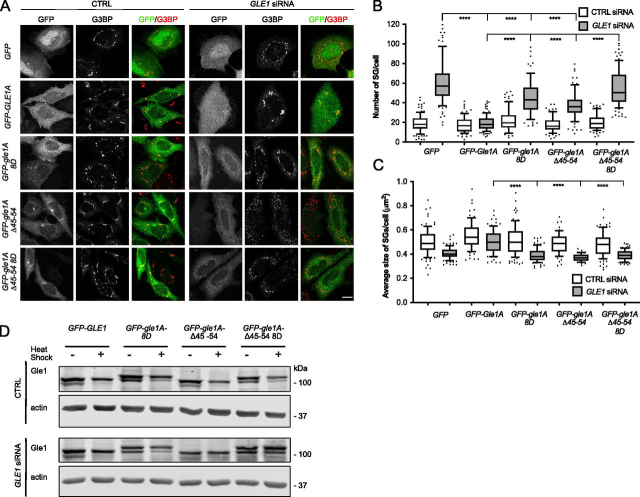
Gle1 is a conserved, essential regulator of DEAD-box RNA helicases, with critical roles defined in mRNA export, translation initiation, translation termination, and stress granule formation. Mechanisms that specify which, where, and when DDXs are targeted by Gle1 are critical to understand. In addition to roles for stress-induced phosphorylation and inositol hexakisphosphate binding in specifying Gle1 function, Gle1 oligomerizes via its N-terminal domain in a phosphorylation-dependent manner. However, a thorough analysis of the role for Gle1 self-association is lacking. Here, we find that Gle1 self-association is driven by two distinct regions: a coiled-coil domain and a novel 10-amino acid aggregation-prone region, both of which are necessary for proper Gle1 oligomerization. By exogenous expression in HeLa cells, we tested the function of a series of mutations that impact the oligomerization domains of the Gle1A and Gle1B isoforms. Gle1 oligomerization is necessary for many, but not all aspects of Gle1A and Gle1B function, and the requirements for each interaction domain differ. Whereas the coiled-coil domain and aggregation-prone region additively contribute to competent mRNA export and stress granule formation, both self-association domains are independently required for regulation of translation under cellular stress. In contrast, Gle1 self-association is dispensable for phosphorylation and nonstressed translation initiation. Collectively, we reveal self-association functions as an additional mode of Gle1 regulation to ensure proper mRNA export and translation. This work also provides further insight into the mechanisms underlying human gle1 disease mutants found in prenatally lethal forms of arthrogryposis.
Keywords: DEAD-box protein; Gle1; mRNA; mRNA export; nuclear pore; oligomerization; protein aggregation; stress granules; translation regulation.
© 2020 Mason and Wente.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures

Similar articles
-
MAPK- and glycogen synthase kinase 3-mediated phosphorylation regulates the DEAD-box protein modulator Gle1 for control of stress granule dynamics.J Biol Chem. 2019 Jan 11;294(2):559-575. doi: 10.1074/jbc.RA118.005749. Epub 2018 Nov 14. J Biol Chem. 2019. PMID: 30429220 Free PMC article.
-
The mRNA export factor Gle1 and inositol hexakisphosphate regulate distinct stages of translation.Cell. 2008 Aug 22;134(4):624-33. doi: 10.1016/j.cell.2008.06.027. Cell. 2008. PMID: 18724935 Free PMC article.
-
Gle1 functions during mRNA export in an oligomeric complex that is altered in human disease.Cell. 2013 Oct 24;155(3):582-93. doi: 10.1016/j.cell.2013.09.023. Epub 2013 Oct 24. Cell. 2013. PMID: 24243016 Free PMC article.
-
Insights into mRNA export-linked molecular mechanisms of human disease through a Gle1 structure-function analysis.Adv Biol Regul. 2014 Jan;54:74-91. doi: 10.1016/j.jbior.2013.10.002. Epub 2013 Nov 13. Adv Biol Regul. 2014. PMID: 24275432 Free PMC article. Review.
-
Dbp5, Gle1-IP6 and Nup159: a working model for mRNP export.Nucleus. 2011 Nov-Dec;2(6):540-8. doi: 10.4161/nucl.2.6.17881. Epub 2011 Nov 1. Nucleus. 2011. PMID: 22064466 Free PMC article. Review.
Cited by
-
A New Phase of Networking: The Molecular Composition and Regulatory Dynamics of Mammalian Stress Granules.Chem Rev. 2023 Jul 26;123(14):9036-9064. doi: 10.1021/acs.chemrev.2c00608. Epub 2023 Jan 20. Chem Rev. 2023. PMID: 36662637 Free PMC article. Review.
-
Gle1 is required for tRNA to stimulate Dbp5 ATPase activity in vitro and to promote Dbp5 mediated tRNA export in vivo.bioRxiv [Preprint]. 2023 Nov 9:2023.06.29.547072. doi: 10.1101/2023.06.29.547072. bioRxiv. 2023. Update in: Elife. 2024 Jan 08;12:RP89835. doi: 10.7554/eLife.89835 PMID: 37425677 Free PMC article. Updated. Preprint.
-
Proteomic Profiles and Protein Network Analysis of Primary Human Leukocytes Revealed Possible Clearance Biomarkers for Staphylococcus aureus Infection.Curr Microbiol. 2023 Sep 4;80(10):335. doi: 10.1007/s00284-023-03450-6. Curr Microbiol. 2023. PMID: 37665379
-
Advances in the understanding of nuclear pore complexes in human diseases.J Cancer Res Clin Oncol. 2024 Jul 30;150(7):374. doi: 10.1007/s00432-024-05881-5. J Cancer Res Clin Oncol. 2024. PMID: 39080077 Free PMC article. Review.
-
Gle1 is required for tRNA to stimulate Dbp5 ATPase activity in vitro and promote Dbp5-mediated tRNA export in vivo in Saccharomyces cerevisiae.Elife. 2024 Jan 8;12:RP89835. doi: 10.7554/eLife.89835. Elife. 2024. PMID: 38189406 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
