

Delineation of a molecularly distinct terminally differentiated memory CD8 T cell population
- PMID: 32978300
- PMCID: PMC7568335
- DOI: 10.1073/pnas.2008571117
Delineation of a molecularly distinct terminally differentiated memory CD8 T cell population
Abstract
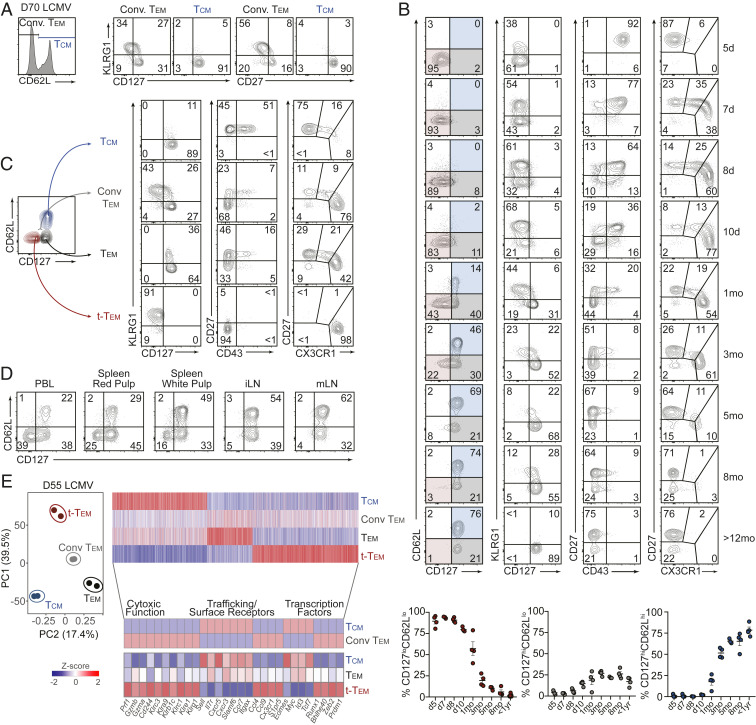
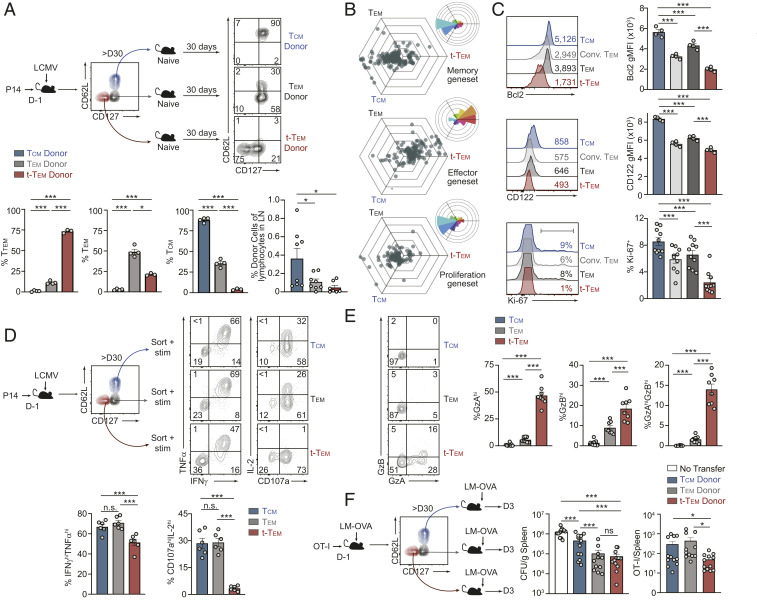
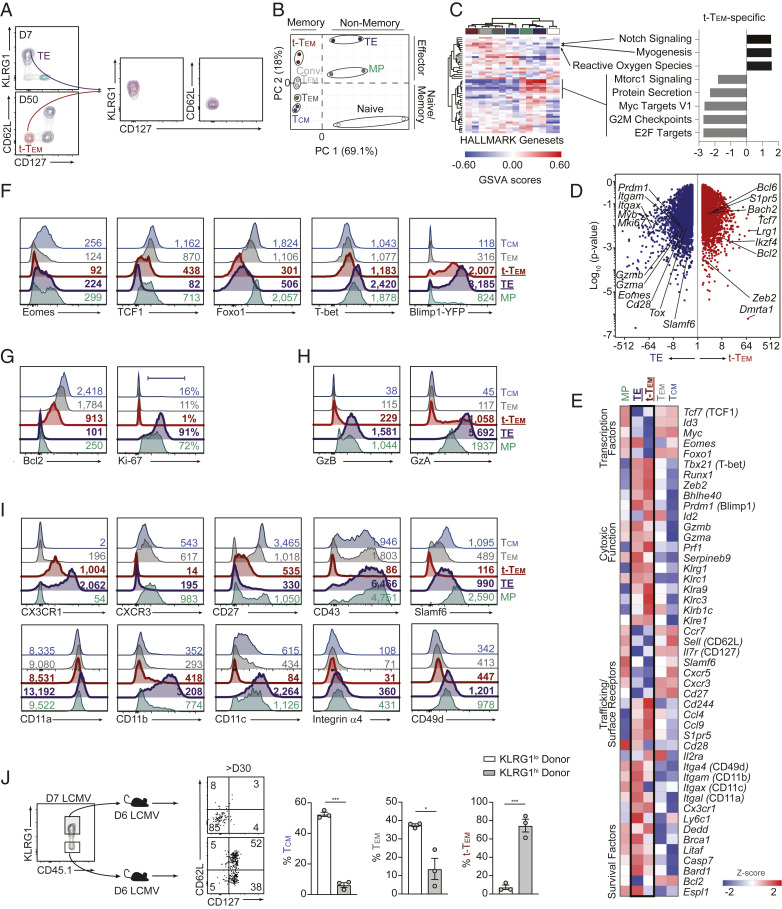
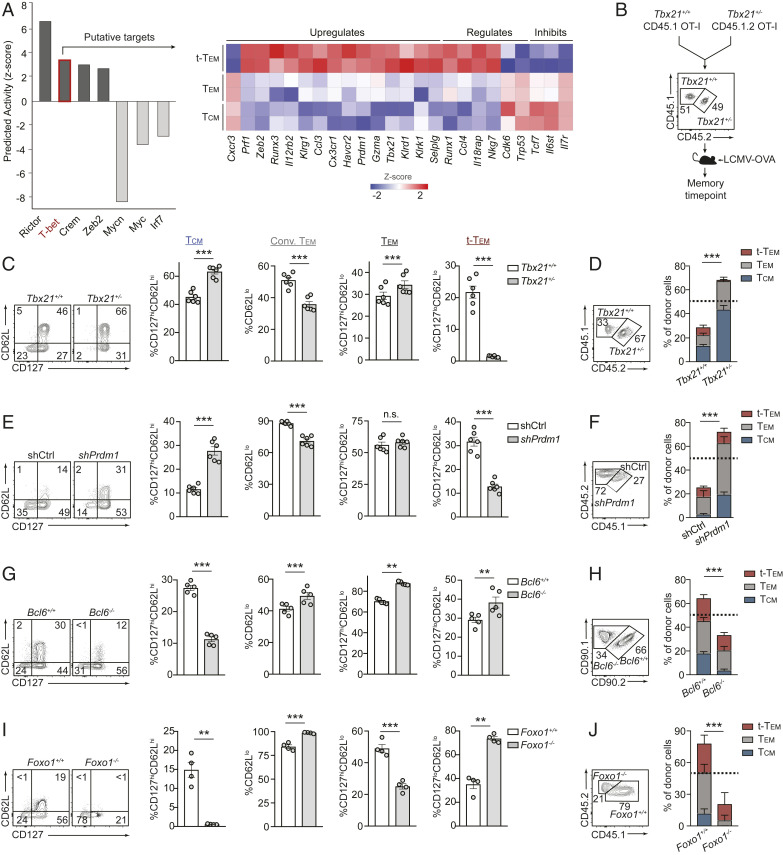
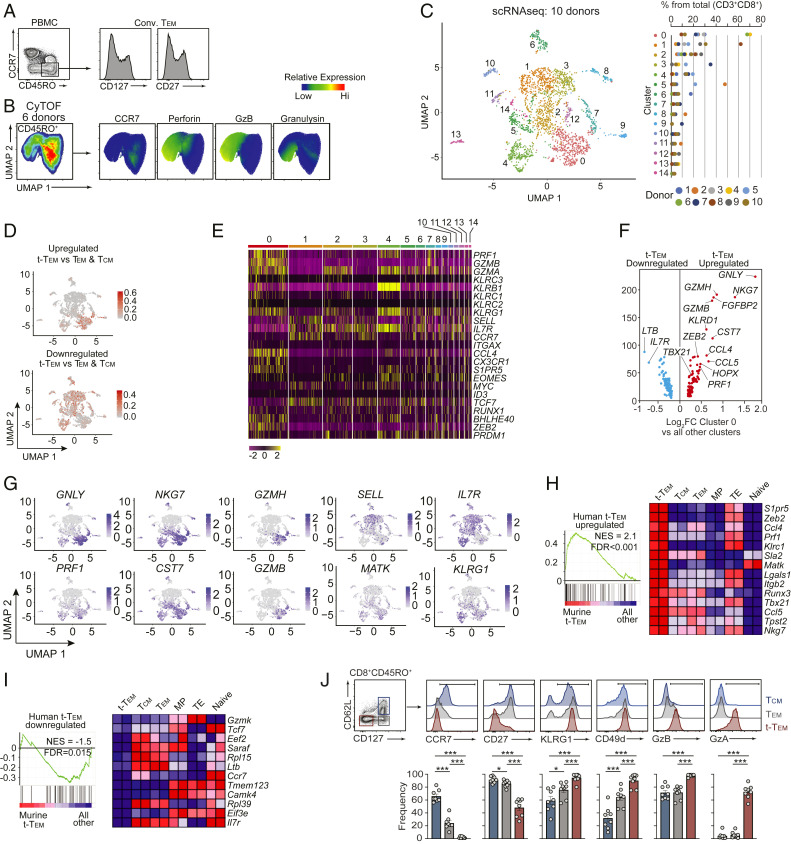
Memory CD8 T cells provide durable protection against diverse intracellular pathogens and can be broadly segregated into distinct circulating and tissue-resident populations. Paradigmatic studies have demonstrated that circulating memory cells can be further divided into effector memory (Tem) and central memory (Tcm) populations based on discrete functional characteristics. Following resolution of infection, we identified a persisting antigen-specific CD8 T cell population that was terminally fated with potent effector function but maintained memory T cell qualities and conferred robust protection against reinfection. Notably, this terminally differentiated effector memory CD8 T cell population (terminal-Tem) was conflated within the conventional Tem population, prompting redefinition of the classical characteristics of Tem cells. Murine terminal-Tem were transcriptionally, functionally, and developmentally unique compared to Tem cells. Through mass cytometry and single-cell RNA sequencing (RNA-seq) analyses of human peripheral blood from healthy individuals, we also identified an analogous terminal-Tem population of CD8 T cells that was transcriptionally distinct from Tem and Tcm Key findings from this study show that parsing of terminal-Tem from conventionally defined Tem challenge the reported characteristics of Tem biology, including enhanced presence in lymphoid tissues, robust IL-2 production, and recall potential, greater than expected homeostatic fitness, refined transcription factor dependencies, and a distinct molecular phenotype. Classification of terminal-Tem and clarification of Tem biology hold broad implications for understanding the molecular regulation of memory cell states and harnessing immunological memory to improve immunotherapies.
Keywords: T cells; immunology; infection; memory T cells.
Conflict of interest statement
Competing interest statement: A.W.G. serves on the scientific advisory boards of Pandion Therapeutics and Arsenal Bio.
Figures
Similar articles
-
Circulating memory CD8+ T cells are limited in forming CD103+ tissue-resident memory T cells at mucosal sites after reinfection.Eur J Immunol. 2021 Jan;51(1):151-166. doi: 10.1002/eji.202048737. Epub 2020 Aug 31. Eur J Immunol. 2021. PMID: 32762051
-
The major CD8 T cell effector memory subset in the normal and Chlamydia trachomatis-infected human endocervix is low in perforin.BMC Immunol. 2012 Dec 7;13:66. doi: 10.1186/1471-2172-13-66. BMC Immunol. 2012. PMID: 23216954 Free PMC article.
-
IL-15-Independent Maintenance of Tissue-Resident and Boosted Effector Memory CD8 T Cells.J Immunol. 2016 May 1;196(9):3920-6. doi: 10.4049/jimmunol.1502337. Epub 2016 Mar 21. J Immunol. 2016. PMID: 27001957 Free PMC article.
-
Genealogy, Dendritic Cell Priming, and Differentiation of Tissue-Resident Memory CD8+ T Cells.Front Immunol. 2018 Jul 31;9:1751. doi: 10.3389/fimmu.2018.01751. eCollection 2018. Front Immunol. 2018. PMID: 30108585 Free PMC article. Review.
-
Area under Immunosurveillance: Dedicated Roles of Memory CD8 T-Cell Subsets.Cold Spring Harb Perspect Biol. 2020 Nov 2;12(11):a037796. doi: 10.1101/cshperspect.a037796. Cold Spring Harb Perspect Biol. 2020. PMID: 32839203 Free PMC article. Review.
Cited by
-
Hypoxia-inducible factor activity promotes antitumor effector function and tissue residency by CD8+ T cells.J Clin Invest. 2021 Apr 1;131(7):e143729. doi: 10.1172/JCI143729. J Clin Invest. 2021. PMID: 33792560 Free PMC article.
-
CTLs Get SMAD When Pathogens Tell Them Where to Go.J Immunol. 2022 Sep 15;209(6):1025-1032. doi: 10.4049/jimmunol.2200345. J Immunol. 2022. PMID: 36130123 Free PMC article. Review.
-
Pregnancy imparts distinct systemic adaptive immune function.Am J Reprod Immunol. 2022 Nov;88(5):e13606. doi: 10.1111/aji.13606. Epub 2022 Sep 6. Am J Reprod Immunol. 2022. PMID: 35989229 Free PMC article.
-
PI3Kδ coordinates transcriptional, chromatin, and metabolic changes to promote effector CD8+ T cells at the expense of central memory.Cell Rep. 2021 Oct 12;37(2):109804. doi: 10.1016/j.celrep.2021.109804. Cell Rep. 2021. PMID: 34644563 Free PMC article.
-
HMGB2 regulates the differentiation and stemness of exhausted CD8+ T cells during chronic viral infection and cancer.Nat Commun. 2023 Sep 13;14(1):5631. doi: 10.1038/s41467-023-41352-0. Nat Commun. 2023. PMID: 37704621 Free PMC article.
References
-
- Wherry E. J., et al. , Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat. Immunol. 4, 225–234 (2003). - PubMed
-
- Sallusto F., Lenig D., Förster R., Lipp M., Lanzavecchia A., Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401, 708–712 (1999). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
