

Cryo-electron Microscopy Structure, Assembly, and Mechanics Show Morphogenesis and Evolution of Human Picobirnavirus
- PMID: 32938763
- PMCID: PMC7925173
- DOI: 10.1128/JVI.01542-20
Cryo-electron Microscopy Structure, Assembly, and Mechanics Show Morphogenesis and Evolution of Human Picobirnavirus
Abstract
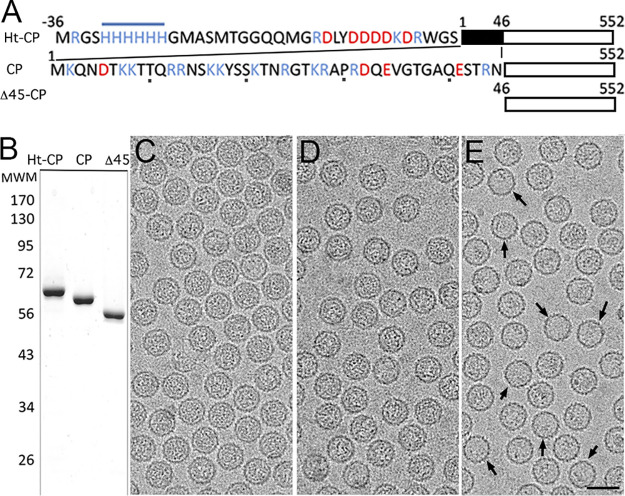
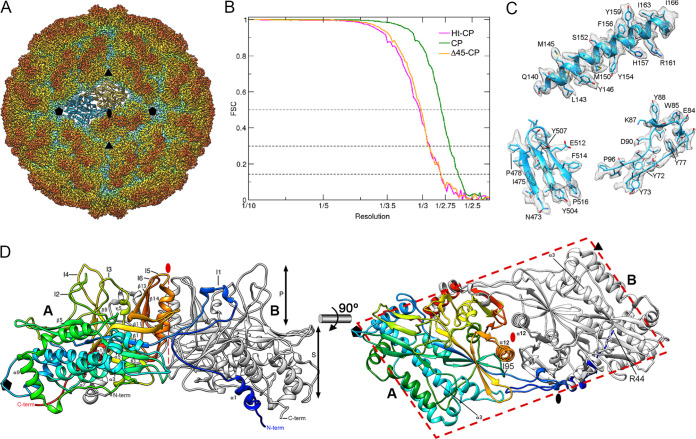
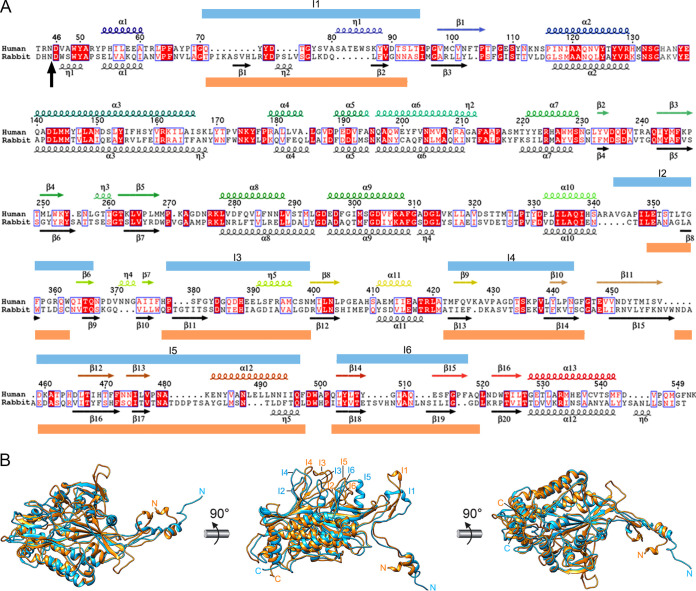
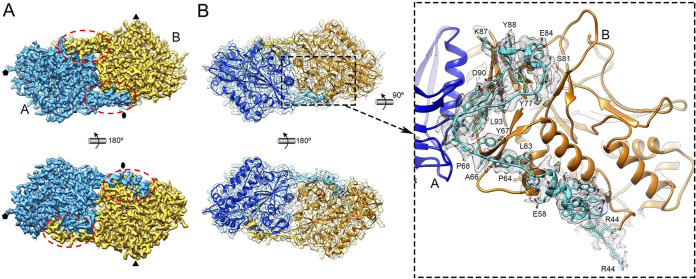
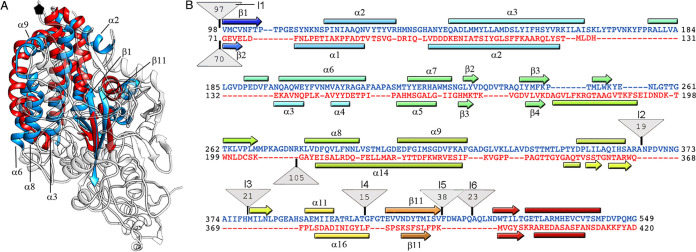
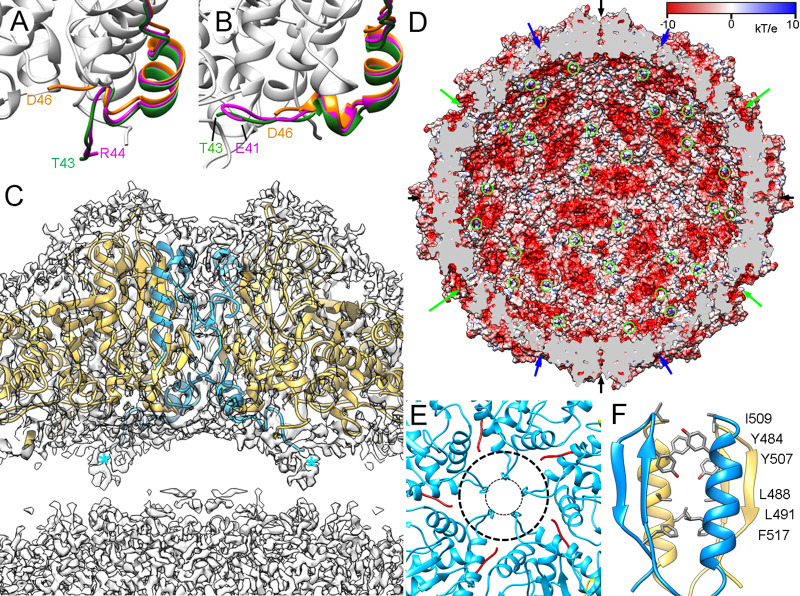
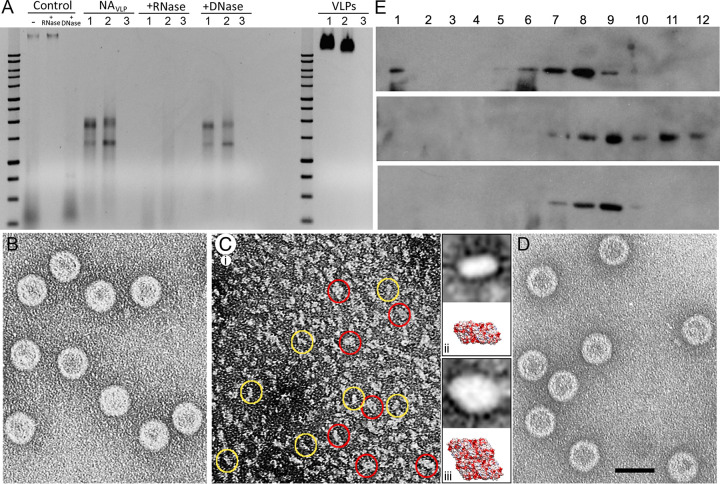
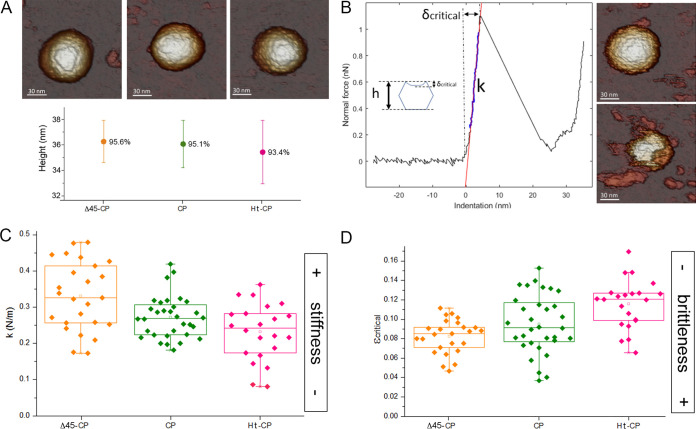
Despite their diversity, most double-stranded-RNA (dsRNA) viruses share a specialized T=1 capsid built from dimers of a single protein that provides a platform for genome transcription and replication. This ubiquitous capsid remains structurally undisturbed throughout the viral cycle, isolating the genome to avoid triggering host defense mechanisms. Human picobirnavirus (hPBV) is a dsRNA virus frequently associated with gastroenteritis, although its pathogenicity is yet undefined. Here, we report the cryo-electron microscopy (cryo-EM) structure of hPBV at 2.6-Å resolution. The capsid protein (CP) is arranged in a single-shelled, ∼380-Å-diameter T=1 capsid with a rough outer surface similar to that of dsRNA mycoviruses. The hPBV capsid is built of 60 quasisymmetric CP dimers (A and B) stabilized by domain swapping, and only the CP-A N-terminal basic region interacts with the packaged nucleic acids. hPBV CP has an α-helical domain with a fold similar to that of fungal partitivirus CP, with many domain insertions in its C-terminal half. In contrast to dsRNA mycoviruses, hPBV has an extracellular life cycle phase like complex reoviruses, which indicates that its own CP probably participates in cell entry. Using an in vitro reversible assembly/disassembly system of hPBV, we isolated tetramers as possible assembly intermediates. We used atomic force microscopy to characterize the biophysical properties of hPBV capsids with different cargos (host nucleic acids or proteins) and found that the CP N-terminal segment not only is involved in nucleic acid interaction/packaging but also modulates the mechanical behavior of the capsid in conjunction with the cargo.IMPORTANCE Despite intensive study, human virus sampling is still sparse, especially for viruses that cause mild or asymptomatic disease. Human picobirnavirus (hPBV) is a double-stranded-RNA virus, broadly dispersed in the human population, but its pathogenicity is uncertain. Here, we report the hPBV structure derived from cryo-electron microscopy (cryo-EM) and reconstruction methods using three capsid protein variants (of different lengths and N-terminal amino acid compositions) that assemble as virus-like particles with distinct properties. The hPBV near-atomic structure reveals a quasisymmetric dimer as the structural subunit and tetramers as possible assembly intermediates that coassemble with nucleic acids. Our structural studies and atomic force microscopy analyses indicate that hPBV capsids are potentially excellent nanocages for gene therapy and targeted drug delivery in humans.
Keywords: 3D cryo-EM; capsid protein; dsRNA virus; hPBV; virus assembly.
Copyright © 2020 American Society for Microbiology.
Figures
Similar articles
-
Heterodimers as the Structural Unit of the T=1 Capsid of the Fungal Double-Stranded RNA Rosellinia necatrix Quadrivirus 1.J Virol. 2016 Nov 28;90(24):11220-11230. doi: 10.1128/JVI.01013-16. Print 2016 Dec 15. J Virol. 2016. PMID: 27707923 Free PMC article.
-
Structure and assembly of double-stranded RNA mycoviruses.Adv Virus Res. 2020;108:213-247. doi: 10.1016/bs.aivir.2020.08.001. Epub 2020 Sep 16. Adv Virus Res. 2020. PMID: 33837717
-
Acquisition of functions on the outer capsid surface during evolution of double-stranded RNA fungal viruses.PLoS Pathog. 2017 Dec 8;13(12):e1006755. doi: 10.1371/journal.ppat.1006755. eCollection 2017 Dec. PLoS Pathog. 2017. PMID: 29220409 Free PMC article.
-
Cryo-electron microscopy for the study of virus assembly.Nat Chem Biol. 2020 Mar;16(3):231-239. doi: 10.1038/s41589-020-0477-1. Epub 2020 Feb 20. Nat Chem Biol. 2020. PMID: 32080621 Review.
-
Capsid Structure of dsRNA Fungal Viruses.Viruses. 2018 Sep 7;10(9):481. doi: 10.3390/v10090481. Viruses. 2018. PMID: 30205532 Free PMC article. Review.
Cited by
-
The structure of a plant-specific partitivirus capsid reveals a unique coat protein domain architecture with an intrinsically disordered protrusion.Commun Biol. 2021 Oct 6;4(1):1155. doi: 10.1038/s42003-021-02687-w. Commun Biol. 2021. PMID: 34615994 Free PMC article.
-
Atomic Structure of the Trichomonas vaginalis Double-Stranded RNA Virus 2.mBio. 2021 Mar 30;12(2):e02924-20. doi: 10.1128/mBio.02924-20. mBio. 2021. PMID: 33785622 Free PMC article.
-
Capsid structure of a fungal dsRNA megabirnavirus reveals its previously unidentified surface architecture.PLoS Pathog. 2023 Feb 27;19(2):e1011162. doi: 10.1371/journal.ppat.1011162. eCollection 2023 Feb. PLoS Pathog. 2023. PMID: 36848381 Free PMC article.
-
High-resolution comparative atomic structures of two Giardiavirus prototypes infecting G. duodenalis parasite.PLoS Pathog. 2024 Apr 10;20(4):e1012140. doi: 10.1371/journal.ppat.1012140. eCollection 2024 Apr. PLoS Pathog. 2024. PMID: 38598600 Free PMC article.
References
-
- Patton JT. 2008. Segmented double-stranded RNA viruses. Structure and molecular biology. Caister Academic Press, Norfolk, United Kingdom.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
