

Antiviral Activity of Type I, II, and III Interferons Counterbalances ACE2 Inducibility and Restricts SARS-CoV-2
- PMID: 32913009
- PMCID: PMC7484541
- DOI: 10.1128/mBio.01928-20
Antiviral Activity of Type I, II, and III Interferons Counterbalances ACE2 Inducibility and Restricts SARS-CoV-2
Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the cause of coronavirus disease 2019 (COVID-19), is a recently emerged respiratory coronavirus that has infected >23 million people worldwide with >800,000 deaths. Few COVID-19 therapeutics are available, and the basis for severe infections is poorly understood. Here, we investigated properties of type I (β), II (γ), and III (λ1) interferons (IFNs), potent immune cytokines that are normally produced during infection and that upregulate IFN-stimulated gene (ISG) effectors to limit virus replication. IFNs are already in clinical trials to treat COVID-19. However, recent studies highlight the potential for IFNs to enhance expression of host angiotensin-converting enzyme 2 (ACE2), suggesting that IFN therapy or natural coinfections could exacerbate COVID-19 by upregulating this critical virus entry receptor. Using a cell line model, we found that beta interferon (IFN-β) strongly upregulated expression of canonical antiviral ISGs, as well as ACE2 at the mRNA and cell surface protein levels. Strikingly, IFN-λ1 upregulated antiviral ISGs, but ACE2 mRNA was only marginally elevated and did not lead to detectably increased ACE2 protein at the cell surface. IFN-γ induced the weakest ISG response but clearly enhanced surface expression of ACE2. Importantly, all IFN types inhibited SARS-CoV-2 replication in a dose-dependent manner, and IFN-β and IFN-λ1 exhibited potent antiviral activity in primary human bronchial epithelial cells. Our data imply that type-specific mechanisms or kinetics shape IFN-enhanced ACE2 transcript and cell surface levels but that the antiviral action of IFNs against SARS-CoV-2 counterbalances any proviral effects of ACE2 induction. These insights should aid in evaluating the benefits of specific IFNs, particularly IFN-λ, as repurposed therapeutics.IMPORTANCE Repurposing existing, clinically approved, antiviral drugs as COVID-19 therapeutics is a rapid way to help combat the SARS-CoV-2 pandemic. Interferons (IFNs) usually form part of the body's natural innate immune defenses against viruses, and they have been used with partial success to treat previous new viral threats, such as HIV, hepatitis C virus, and Ebola virus. Nevertheless, IFNs can have undesirable side effects, and recent reports indicate that IFNs upregulate the expression of host ACE2 (a critical entry receptor for SARS-CoV-2), raising the possibility that IFN treatments could exacerbate COVID-19. Here, we studied the antiviral- and ACE2-inducing properties of different IFN types in both a human lung cell line model and primary human bronchial epithelial cells. We observed differences between IFNs with respect to their induction of antiviral genes and abilities to enhance the cell surface expression of ACE2. Nevertheless, all the IFNs limited SARS-CoV-2 replication, suggesting that their antiviral actions can counterbalance increased ACE2.
Keywords: ACE2; COVID-19; SARS-CoV-2; coronavirus; immunotherapy; interferons; receptors.
Copyright © 2020 Busnadiego et al.
Figures
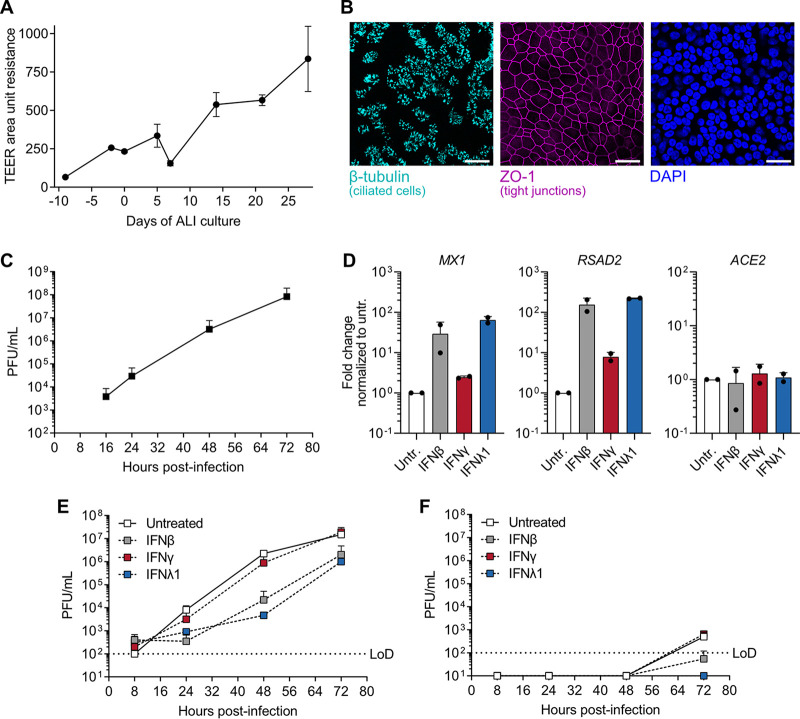
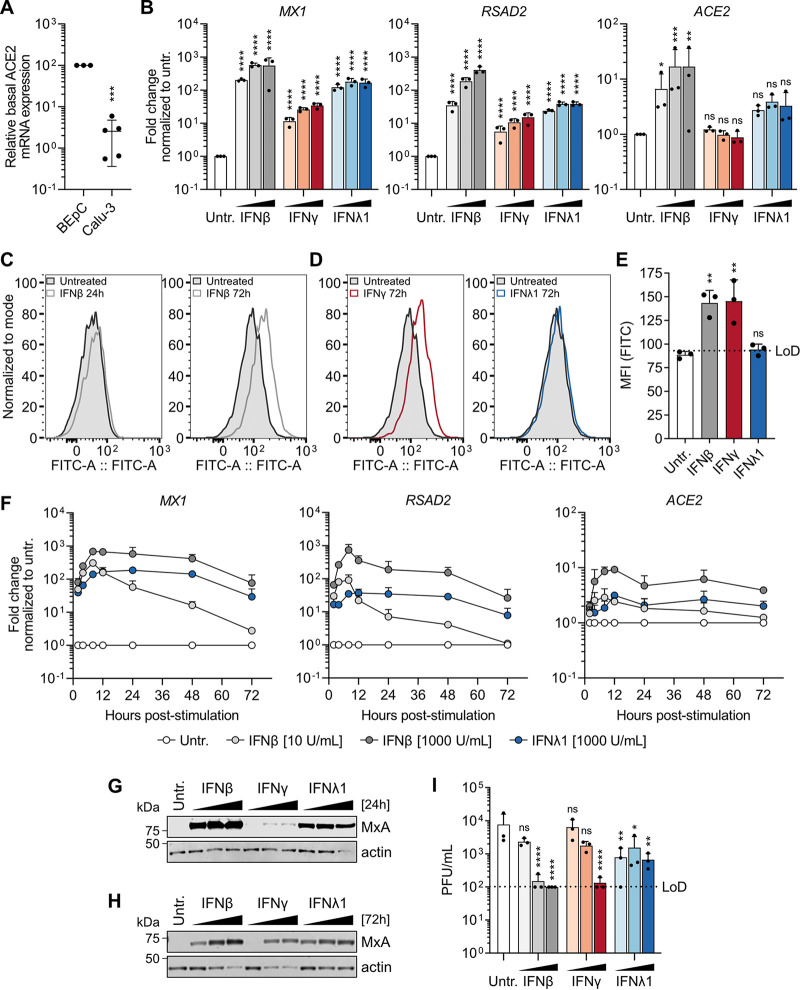
Similar articles
-
Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures.J Virol. 2020 Sep 15;94(19):e00985-20. doi: 10.1128/JVI.00985-20. Print 2020 Sep 15. J Virol. 2020. PMID: 32699094 Free PMC article.
-
Inhibition of SARS-CoV-2 by type I and type III interferons.J Biol Chem. 2020 Oct 9;295(41):13958-13964. doi: 10.1074/jbc.AC120.013788. Epub 2020 Jun 25. J Biol Chem. 2020. PMID: 32587093 Free PMC article.
-
A mouse-adapted model of SARS-CoV-2 to test COVID-19 countermeasures.Nature. 2020 Oct;586(7830):560-566. doi: 10.1038/s41586-020-2708-8. Epub 2020 Aug 27. Nature. 2020. PMID: 32854108 Free PMC article.
-
Type I and Type III Interferons - Induction, Signaling, Evasion, and Application to Combat COVID-19.Cell Host Microbe. 2020 Jun 10;27(6):870-878. doi: 10.1016/j.chom.2020.05.008. Epub 2020 May 27. Cell Host Microbe. 2020. PMID: 32464097 Free PMC article. Review.
-
SARS-CoV-2 pandemic and research gaps: Understanding SARS-CoV-2 interaction with the ACE2 receptor and implications for therapy.Theranostics. 2020 Jun 12;10(16):7448-7464. doi: 10.7150/thno.48076. eCollection 2020. Theranostics. 2020. PMID: 32642005 Free PMC article. Review.
Cited by
-
Differentially expressed immune response genes in COVID-19 patients based on disease severity.Aging (Albany NY). 2021 Mar 29;13(7):9265-9276. doi: 10.18632/aging.202877. Epub 2021 Mar 29. Aging (Albany NY). 2021. PMID: 33780352 Free PMC article.
-
Hamster organotypic kidney culture model of early-stage SARS-CoV-2 infection highlights a two-step renal susceptibility.J Tissue Eng. 2022 Sep 6;13:20417314221122130. doi: 10.1177/20417314221122130. eCollection 2022 Jan-Dec. J Tissue Eng. 2022. PMID: 36093433 Free PMC article.
-
Exploring post-SEPSIS and post-COVID-19 syndromes: crossovers from pathophysiology to therapeutic approach.Front Med (Lausanne). 2024 Jan 5;10:1280951. doi: 10.3389/fmed.2023.1280951. eCollection 2023. Front Med (Lausanne). 2024. PMID: 38249978 Free PMC article. Review.
-
Variable susceptibility of intestinal organoid-derived monolayers to SARS-CoV-2 infection.PLoS Biol. 2022 Mar 31;20(3):e3001592. doi: 10.1371/journal.pbio.3001592. eCollection 2022 Mar. PLoS Biol. 2022. PMID: 35358182 Free PMC article.
-
Effectiveness of SARS-CoV-2 Vaccines for Short- and Long-Term Immunity: A General Overview for the Pandemic Contrast.Int J Mol Sci. 2022 Jul 30;23(15):8485. doi: 10.3390/ijms23158485. Int J Mol Sci. 2022. PMID: 35955621 Free PMC article. Review.
References
-
- Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, Zhao X, Huang B, Shi W, Lu R, Niu P, Zhan F, Ma X, Wang D, Xu W, Wu G, Gao GF, Tan W, China Novel Coronavirus Investigating and Research Team. 2020. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med 382:727–733. doi:10.1056/NEJMoa2001017. - DOI - PMC - PubMed
-
- World Health Organization. 2020. Coronavirus disease 19 (COVID-19) situation report. World Health Organization, Geneva, Switzerland.
-
- Hoffmann M, Kleine-Weber H, Schroeder S, Kruger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, Muller MA, Drosten C, Pohlmann S. 2020. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181:271–280.e8. doi:10.1016/j.cell.2020.02.052. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
