

Understanding microcephaly through the study of centrosome regulation in Drosophila neural stem cells
- PMID: 32897294
- PMCID: PMC7606523
- DOI: 10.1042/BST20200261
Understanding microcephaly through the study of centrosome regulation in Drosophila neural stem cells
Abstract
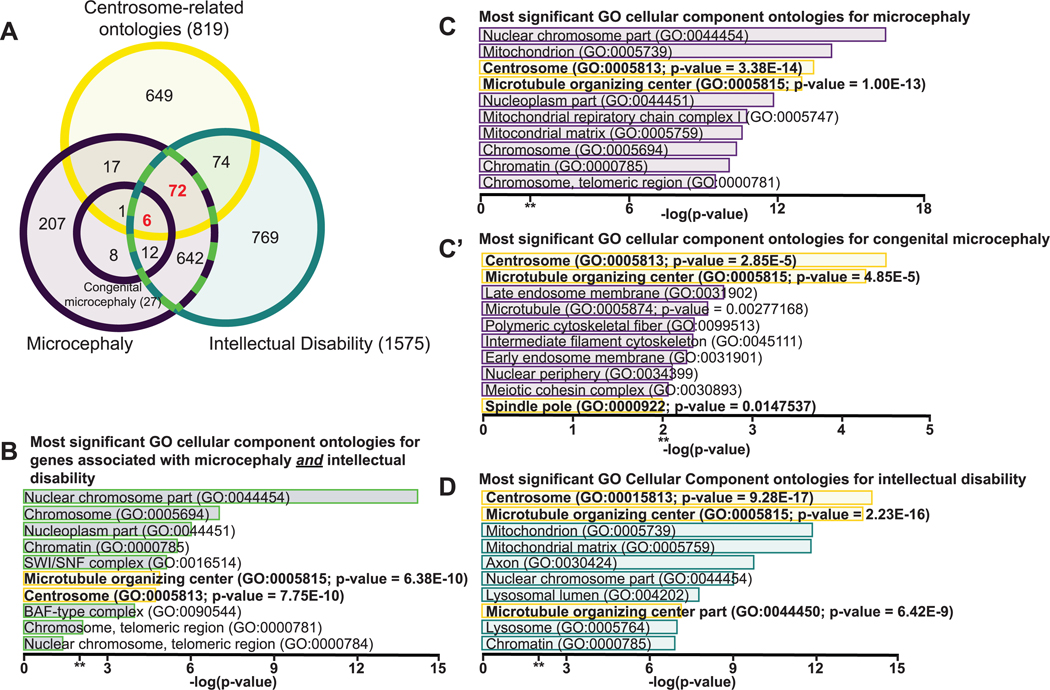
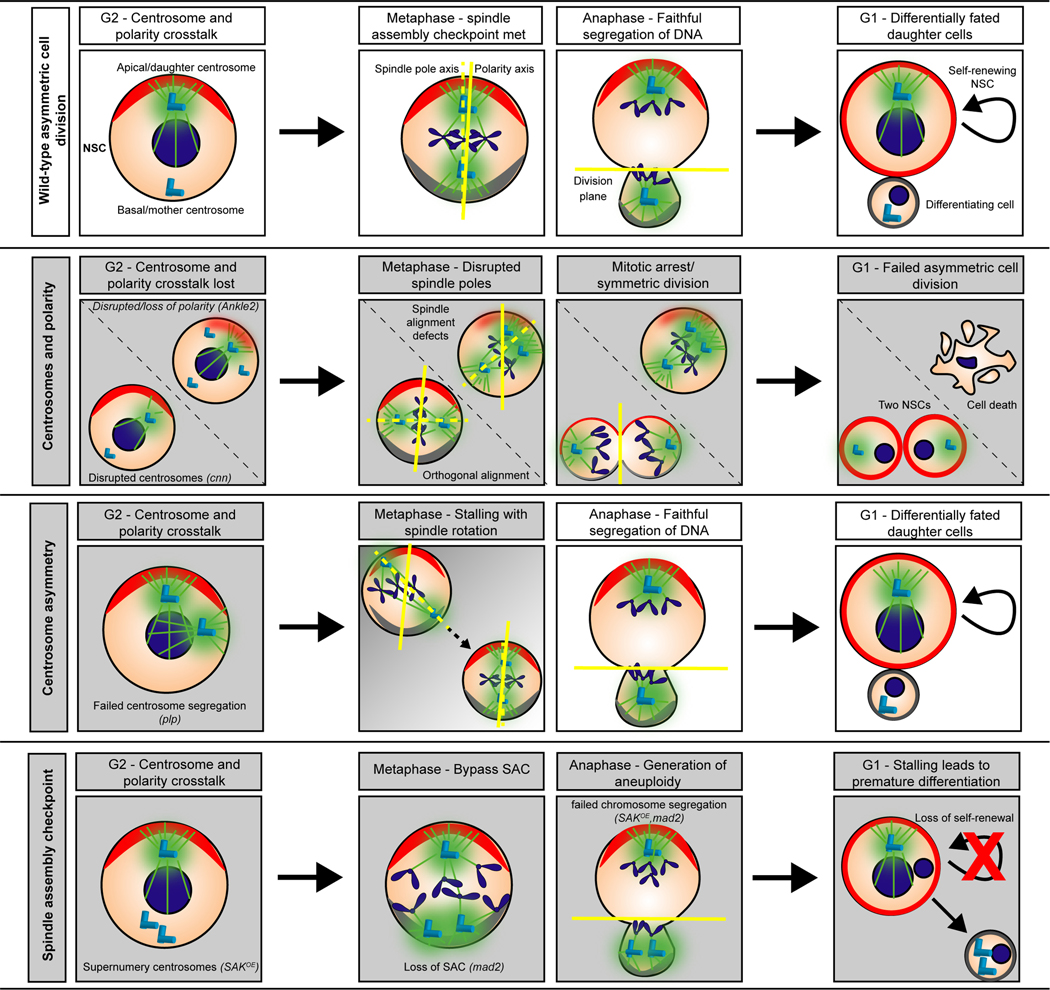
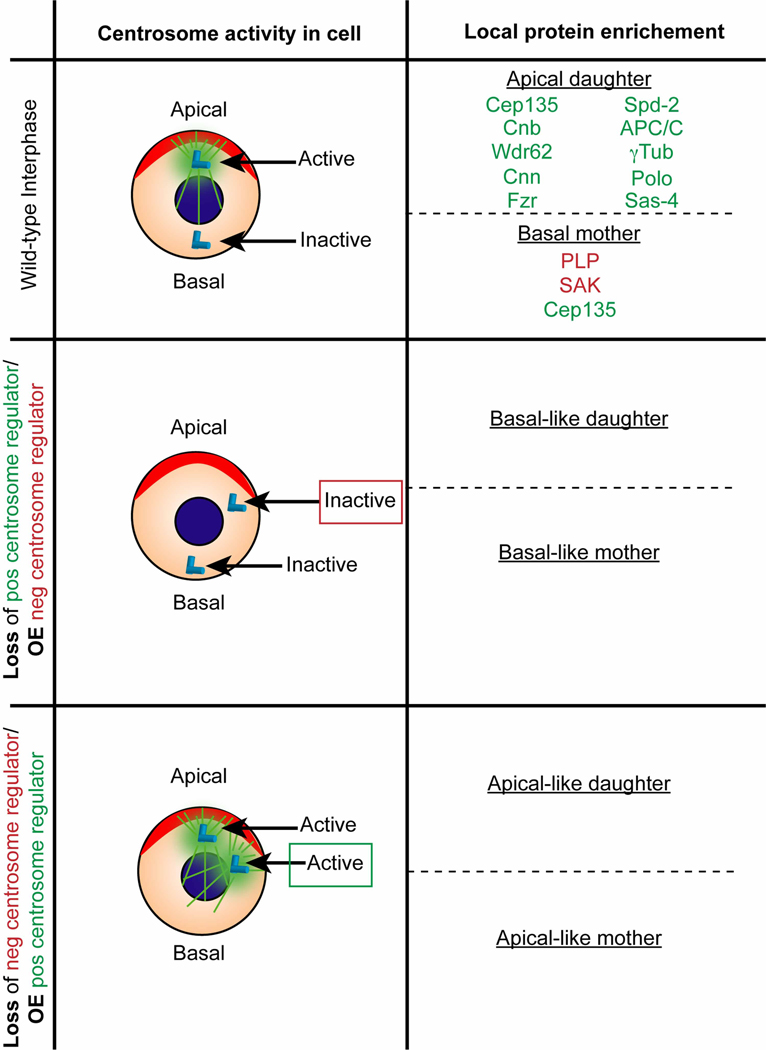
Microcephaly is a rare, yet devastating, neurodevelopmental condition caused by genetic or environmental insults, such as the Zika virus infection. Microcephaly manifests with a severely reduced head circumference. Among the known heritable microcephaly genes, a significant proportion are annotated with centrosome-related ontologies. Centrosomes are microtubule-organizing centers, and they play fundamental roles in the proliferation of the neuronal progenitors, the neural stem cells (NSCs), which undergo repeated rounds of asymmetric cell division to drive neurogenesis and brain development. Many of the genes, pathways, and developmental paradigms that dictate NSC development in humans are conserved in Drosophila melanogaster. As such, studies of Drosophila NSCs lend invaluable insights into centrosome function within NSCs and help inform the pathophysiology of human microcephaly. This mini-review will briefly survey causative links between deregulated centrosome functions and microcephaly with particular emphasis on insights learned from Drosophila NSCs.
Keywords: Drosophila melanogaster; centrosomes; intellectual disability; microcephaly; neural stem cells.
© 2020 The Author(s). Published by Portland Press Limited on behalf of the Biochemical Society.
Figures
Similar articles
-
The Microcephaly-Associated Protein Wdr62/CG7337 Is Required to Maintain Centrosome Asymmetry in Drosophila Neuroblasts.Cell Rep. 2016 Feb 9;14(5):1100-1113. doi: 10.1016/j.celrep.2015.12.097. Epub 2016 Jan 21. Cell Rep. 2016. PMID: 26804909
-
Same but different: pleiotropy in centrosome-related microcephaly.Mol Biol Cell. 2018 Feb 1;29(3):241-246. doi: 10.1091/mbc.E17-03-0192. Mol Biol Cell. 2018. PMID: 29382806 Free PMC article. Review.
-
A centrosomal view of CNS growth.Development. 2018 Nov 6;145(21):dev170613. doi: 10.1242/dev.170613. Development. 2018. PMID: 30401784 Review.
-
A role for a novel centrosome cycle in asymmetric cell division.J Cell Biol. 2007 Apr 9;177(1):13-20. doi: 10.1083/jcb.200612140. Epub 2007 Apr 2. J Cell Biol. 2007. PMID: 17403931 Free PMC article.
-
Centrosome and spindle assembly checkpoint loss leads to neural apoptosis and reduced brain size.J Cell Biol. 2017 May 1;216(5):1255-1265. doi: 10.1083/jcb.201607022. Epub 2017 Mar 28. J Cell Biol. 2017. PMID: 28351851 Free PMC article.
Cited by
-
PCDH12 loss results in premature neuronal differentiation and impeded migration in a cortical organoid model.Cell Rep. 2023 Aug 29;42(8):112845. doi: 10.1016/j.celrep.2023.112845. Epub 2023 Jul 21. Cell Rep. 2023. PMID: 37480564 Free PMC article.
-
Dynamic Changes of Brain Cilia Transcriptomes across the Human Lifespan.Int J Mol Sci. 2021 Sep 27;22(19):10387. doi: 10.3390/ijms221910387. Int J Mol Sci. 2021. PMID: 34638726 Free PMC article.
-
Primary cilia in Parkinson's disease: summative roles in signaling pathways, genes, defective mitochondrial function, and substantia nigra dopaminergic neurons.Front Aging Neurosci. 2024 Sep 18;16:1451655. doi: 10.3389/fnagi.2024.1451655. eCollection 2024. Front Aging Neurosci. 2024. PMID: 39364348 Free PMC article. Review.
-
The international exchange of Drosophila melanogaster strains.Rev Sci Tech. 2022 May;41(1):82-90. doi: 10.20506/rst.41.1.3305. Rev Sci Tech. 2022. PMID: 35925634 Free PMC article.
-
The neurological and non-neurological roles of the primary microcephaly-associated protein ASPM.Front Neurosci. 2023 Aug 3;17:1242448. doi: 10.3389/fnins.2023.1242448. eCollection 2023. Front Neurosci. 2023. PMID: 37599996 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials