

Critical review of non-histone human substrates of metal-dependent lysine deacetylases
- PMID: 32862458
- PMCID: PMC7719617
- DOI: 10.1096/fj.202001301RR
Critical review of non-histone human substrates of metal-dependent lysine deacetylases
Abstract
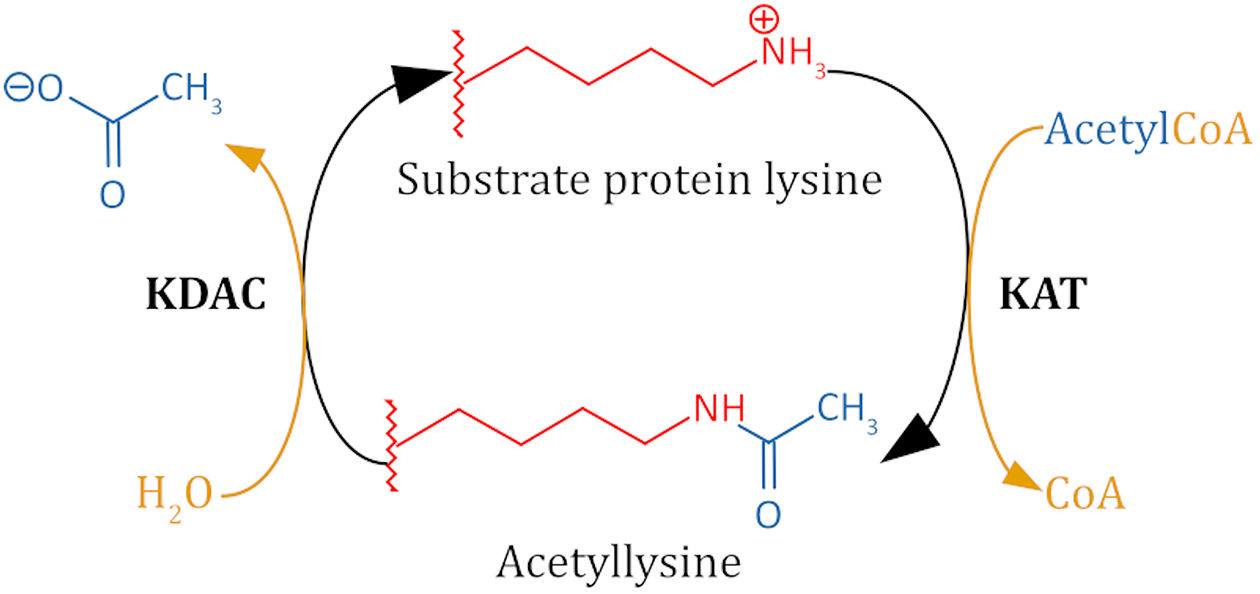
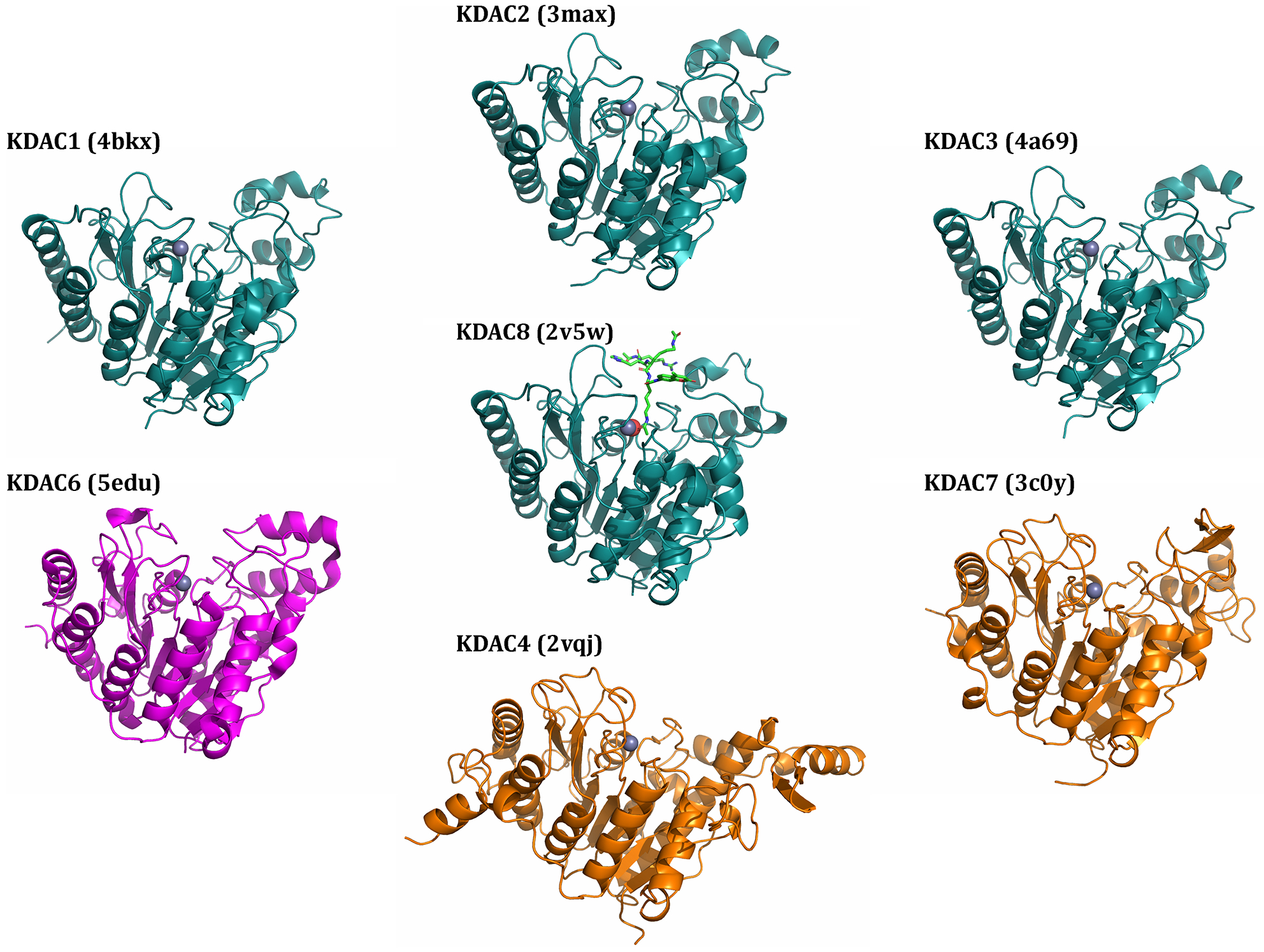
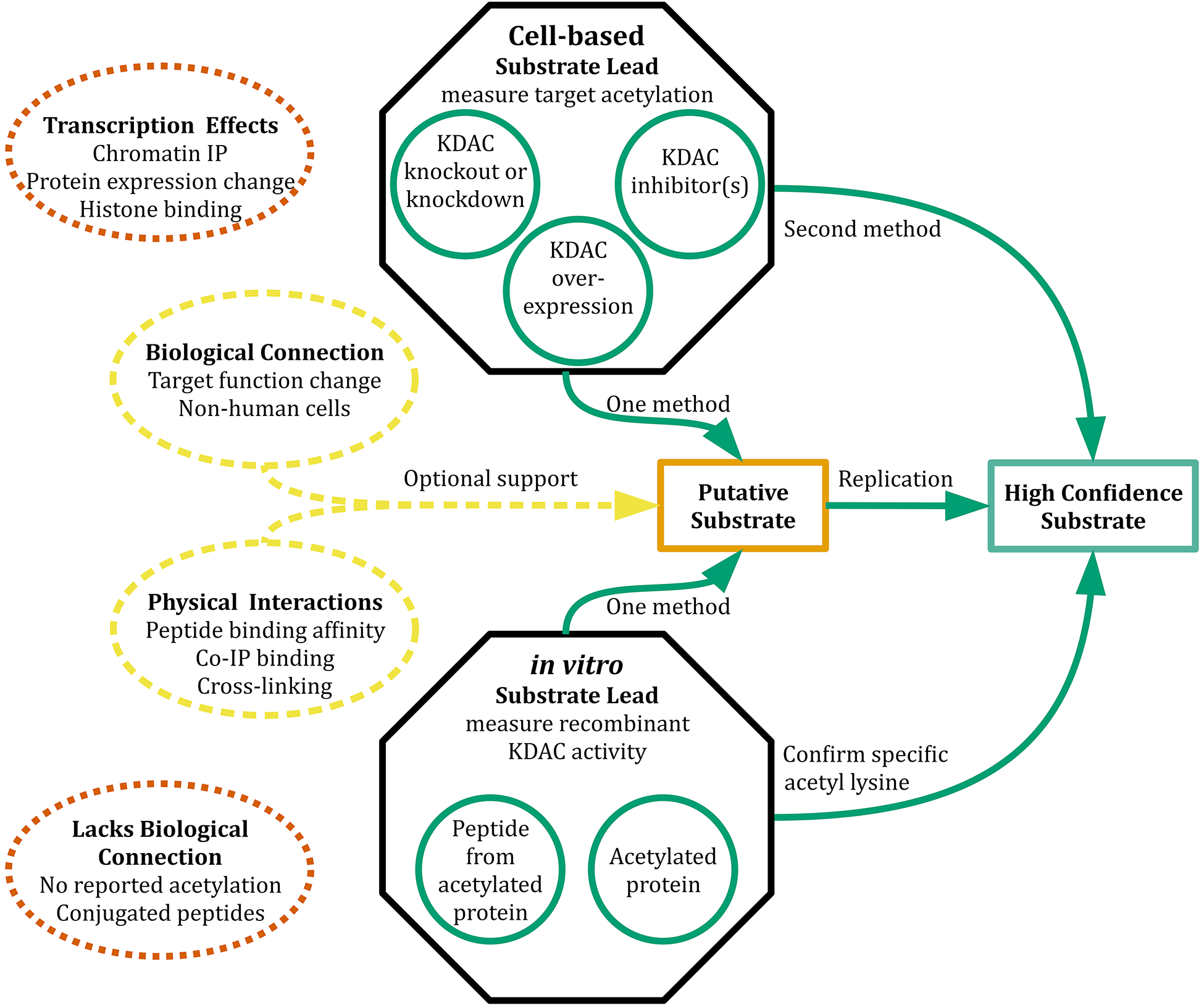
Lysine acetylation is a posttranslational modification that occurs on thousands of human proteins, most of which are cytoplasmic. Acetylated proteins are involved in numerous cellular processes and human diseases. Therefore, how the acetylation/deacetylation cycle is regulated is an important question. Eleven metal-dependent lysine deacetylases (KDACs) have been identified in human cells. These enzymes, along with the sirtuins, are collectively responsible for reversing lysine acetylation. Despite several large-scale studies which have characterized the acetylome, relatively few of the specific acetylated residues have been matched to a proposed KDAC for deacetylation. To understand the function of lysine acetylation, and its association with diseases, specific KDAC-substrate pairs must be identified. Identifying specific substrates of a KDAC is complicated both by the complexity of assaying relevant activity and by the non-catalytic interactions of KDACs with cellular proteins. Here, we discuss in vitro and cell-based experimental strategies used to identify KDAC-substrate pairs and evaluate each for the purpose of directly identifying non-histone substrates of metal-dependent KDACs. We propose criteria for a combination of reproducible experimental approaches that are necessary to establish a direct enzymatic relationship. This critical analysis of the literature identifies 108 proposed non-histone substrate-KDAC pairs for which direct experimental evidence has been reported. Of these, five pairs can be considered well-established, while another thirteen pairs have both cell-based and in vitro evidence but lack independent replication and/or sufficient cell-based evidence. We present a path forward for evaluating the remaining substrate leads and reliably identifying novel KDAC substrates.
Keywords: HDAC; KDAC; histone deacetylases; substrate specificity.
© 2020 Federation of American Societies for Experimental Biology.
Conflict of interest statement
Conflict of Interest Statement
The authors declare that there are no conflicts of interest.
Figures

Similar articles
-
Assays to Study Enzymatic and Non-Enzymatic Protein Lysine Acetylation In Vitro.Curr Protoc. 2021 Nov;1(11):e277. doi: 10.1002/cpz1.277. Curr Protoc. 2021. PMID: 34748287
-
KDAC8 substrate specificity quantified by a biologically relevant, label-free deacetylation assay.Protein Sci. 2015 Dec;24(12):2020-32. doi: 10.1002/pro.2813. Epub 2015 Oct 7. Protein Sci. 2015. PMID: 26402585 Free PMC article.
-
Genome-scale analysis of regulatory protein acetylation enzymes from photosynthetic eukaryotes.BMC Genomics. 2017 Jul 5;18(1):514. doi: 10.1186/s12864-017-3894-0. BMC Genomics. 2017. PMID: 28679357 Free PMC article.
-
Lysine deacetylase (KDAC) regulatory pathways: an alternative approach to selective modulation.ChemMedChem. 2014 Mar;9(3):511-22. doi: 10.1002/cmdc.201300444. Epub 2014 Jan 21. ChemMedChem. 2014. PMID: 24449617 Free PMC article. Review.
-
Building a KATalogue of acetyllysine targeting and function.Brief Funct Genomics. 2016 Mar;15(2):109-18. doi: 10.1093/bfgp/elv045. Epub 2015 Oct 27. Brief Funct Genomics. 2016. PMID: 26512033 Free PMC article. Review.
Cited by
-
Role of Fluorination in the Histone Deacetylase 6 (HDAC6) Selectivity of Benzohydroxamate-Based Inhibitors.ACS Med Chem Lett. 2021 Oct 11;12(11):1810-1817. doi: 10.1021/acsmedchemlett.1c00425. eCollection 2021 Nov 11. ACS Med Chem Lett. 2021. PMID: 34795871 Free PMC article.
-
Lysine Deacetylase Substrate Selectivity: Distinct Interaction Surfaces Drive Positive and Negative Selection for Residues Following Acetyllysine.Biochemistry. 2023 May 2;62(9):1464-1483. doi: 10.1021/acs.biochem.3c00001. Epub 2023 Apr 12. Biochemistry. 2023. PMID: 37043688 Free PMC article.
-
Lysine Deacetylase Substrate Selectivity: A Dynamic Ionic Interaction Specific to KDAC8.Biochemistry. 2021 Aug 24;60(33):2524-2536. doi: 10.1021/acs.biochem.1c00384. Epub 2021 Aug 6. Biochemistry. 2021. PMID: 34357750 Free PMC article.
-
Roles of Histone Deacetylases in Acute Myeloid Leukemia With Fusion Proteins.Front Oncol. 2021 Sep 1;11:741746. doi: 10.3389/fonc.2021.741746. eCollection 2021. Front Oncol. 2021. PMID: 34540702 Free PMC article. Review.
-
Endogenous expression of inactive lysine deacetylases reveals deacetylation-dependent cellular mechanisms.PLoS One. 2023 Sep 18;18(9):e0291779. doi: 10.1371/journal.pone.0291779. eCollection 2023. PLoS One. 2023. PMID: 37721967 Free PMC article.
References
-
- Zhao S, Xu W, Jiang W, Yu W, Lin Y, Zhang T, Yao J, Zhou L, Zeng Y, Li H, Li Y, Shi J, An W, Hancock SM, He F, Qin L, Chin J, Yang P, Chen X, Lei Q, Xiong Y, Guan K-L. Regulation of cellular metabolism by protein lysine acetylation. Science. 2010;327:1000–1004. doi:10.1126/science.1179689 - DOI - PMC - PubMed
-
- Schölz C, Weinert BT, Wagner SA, Beli P, Miyake Y, Qi J, Jensen LJ, Streicher W, McCarthy AR, Westwood NJ, Lain S, Cox J, Matthias P, Mann M, Bradner JE, Choudhary C. Acetylation site specificities of lysine deacetylase inhibitors in human cells. Nat Biotechnol. 2015;33:415–423. doi:10.1038/nbt.3130 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
