

Beyond Trees: Regulons and Regulatory Motif Characterization
- PMID: 32854400
- PMCID: PMC7564462
- DOI: 10.3390/genes11090995
Beyond Trees: Regulons and Regulatory Motif Characterization
Abstract
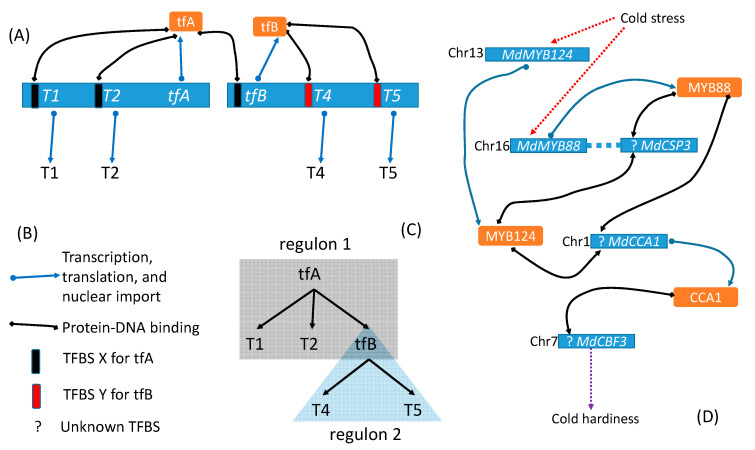
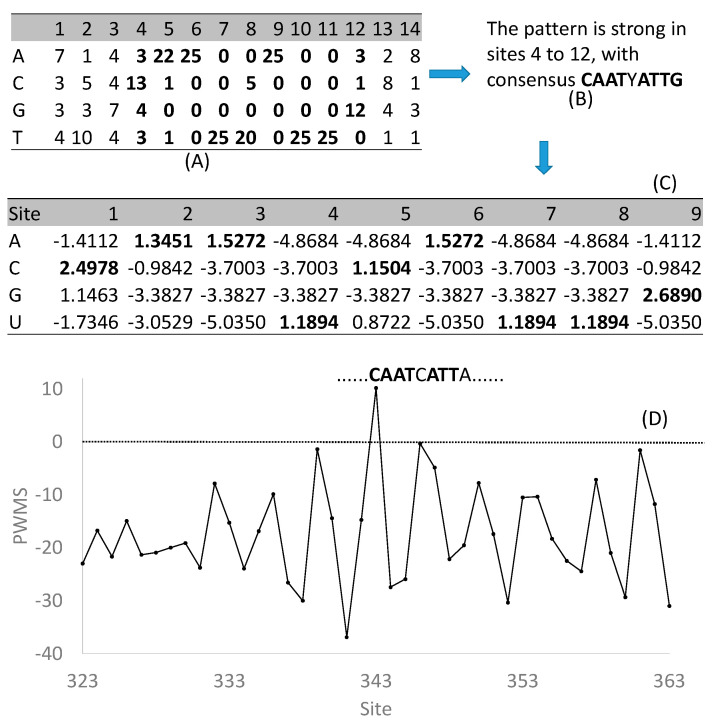
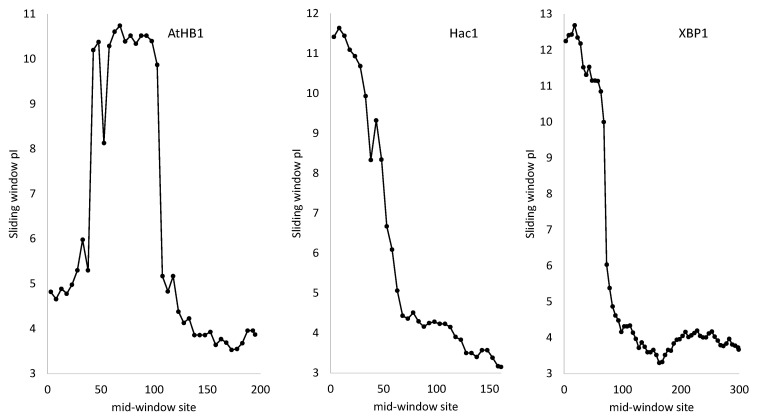
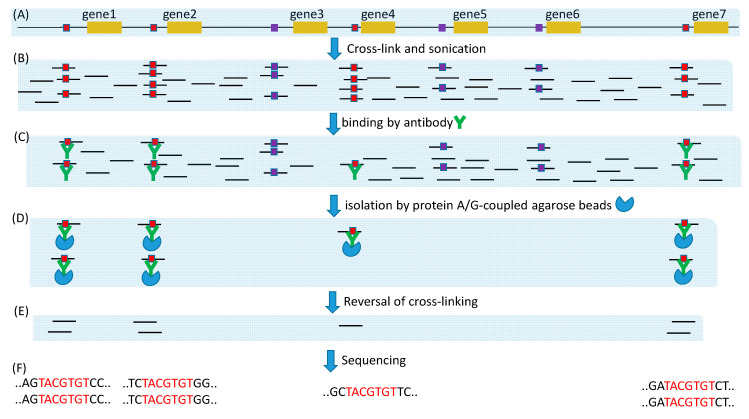
Trees and their seeds regulate their germination, growth, and reproduction in response to environmental stimuli. These stimuli, through signal transduction, trigger transcription factors that alter the expression of various genes leading to the unfolding of the genetic program. A regulon is conceptually defined as a set of target genes regulated by a transcription factor by physically binding to regulatory motifs to accomplish a specific biological function, such as the CO-FT regulon for flowering timing and fall growth cessation in trees. Only with a clear characterization of regulatory motifs, can candidate target genes be experimentally validated, but motif characterization represents the weakest feature of regulon research, especially in tree genetics. I review here relevant experimental and bioinformatics approaches in characterizing transcription factors and their binding sites, outline problems in tree regulon research, and demonstrate how transcription factor databases can be effectively used to aid the characterization of tree regulons.
Keywords: Gibbs sampler; comparative genomics; gene expression; regulatory motifs; regulon; transcription factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Genomic reconstruction of the transcriptional regulatory network in Bacillus subtilis.J Bacteriol. 2013 Jun;195(11):2463-73. doi: 10.1128/JB.00140-13. Epub 2013 Mar 15. J Bacteriol. 2013. PMID: 23504016 Free PMC article.
-
Reconstruction of novel transcription factor regulons through inference of their binding sites.BMC Bioinformatics. 2015 Sep 21;16:299. doi: 10.1186/s12859-015-0685-y. BMC Bioinformatics. 2015. PMID: 26388177 Free PMC article.
-
Comparative genomics of pyridoxal 5'-phosphate-dependent transcription factor regulons in Bacteria.Microb Genom. 2016 Jan 18;2(1):e000047. doi: 10.1099/mgen.0.000047. eCollection 2016 Jan. Microb Genom. 2016. PMID: 28348826 Free PMC article.
-
Regulons of global transcription factors in Corynebacterium glutamicum.Appl Microbiol Biotechnol. 2016 Jan;100(1):45-60. doi: 10.1007/s00253-015-7074-3. Appl Microbiol Biotechnol. 2016. PMID: 26496920 Review.
-
Which came first, the transcriptional regulator or its target genes? An evolutionary perspective into the construction of eukaryotic regulons.Biochim Biophys Acta Gene Regul Mech. 2020 Jun;1863(6):194472. doi: 10.1016/j.bbagrm.2019.194472. Epub 2019 Dec 9. Biochim Biophys Acta Gene Regul Mech. 2020. PMID: 31825805 Review.
Cited by
-
Domains and Functions of Spike Protein in Sars-Cov-2 in the Context of Vaccine Design.Viruses. 2021 Jan 14;13(1):109. doi: 10.3390/v13010109. Viruses. 2021. PMID: 33466921 Free PMC article. Review.
-
Post-Alignment Adjustment and Its Automation.Genes (Basel). 2021 Nov 18;12(11):1809. doi: 10.3390/genes12111809. Genes (Basel). 2021. PMID: 34828415 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
