

Acheron/Larp6 Is a Survival Protein That Protects Skeletal Muscle From Programmed Cell Death During Development
- PMID: 32850788
- PMCID: PMC7405549
- DOI: 10.3389/fcell.2020.00622
Acheron/Larp6 Is a Survival Protein That Protects Skeletal Muscle From Programmed Cell Death During Development
Abstract
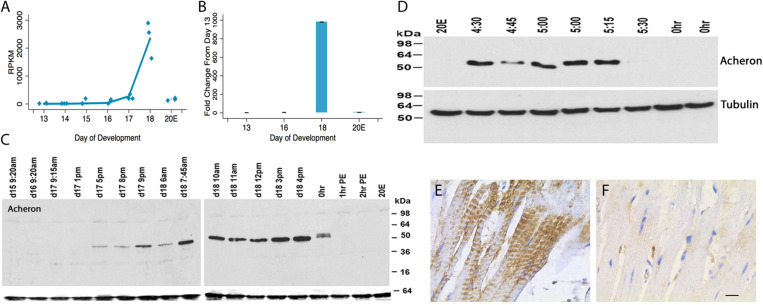
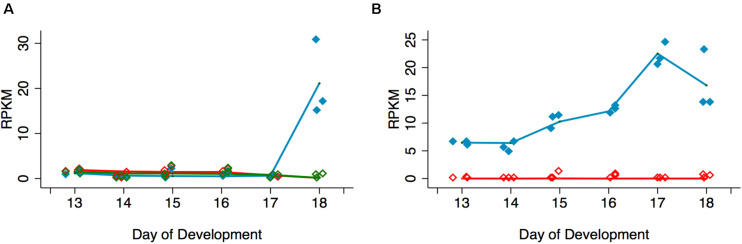
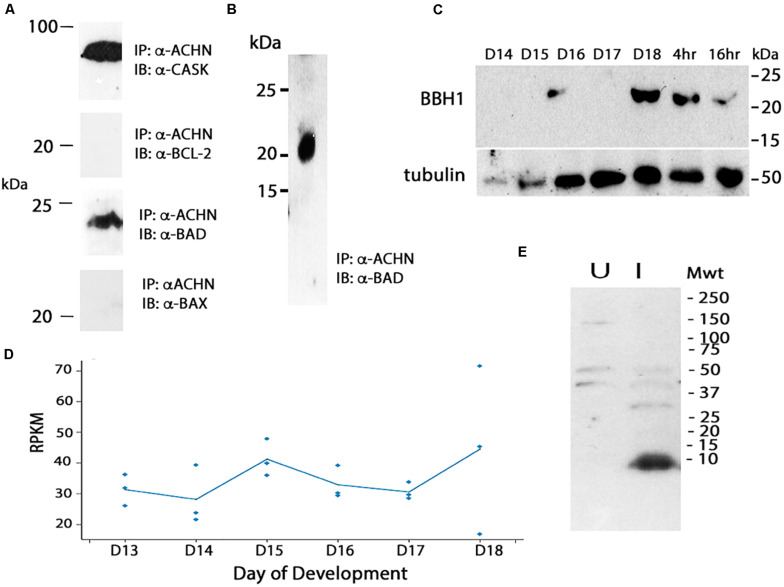
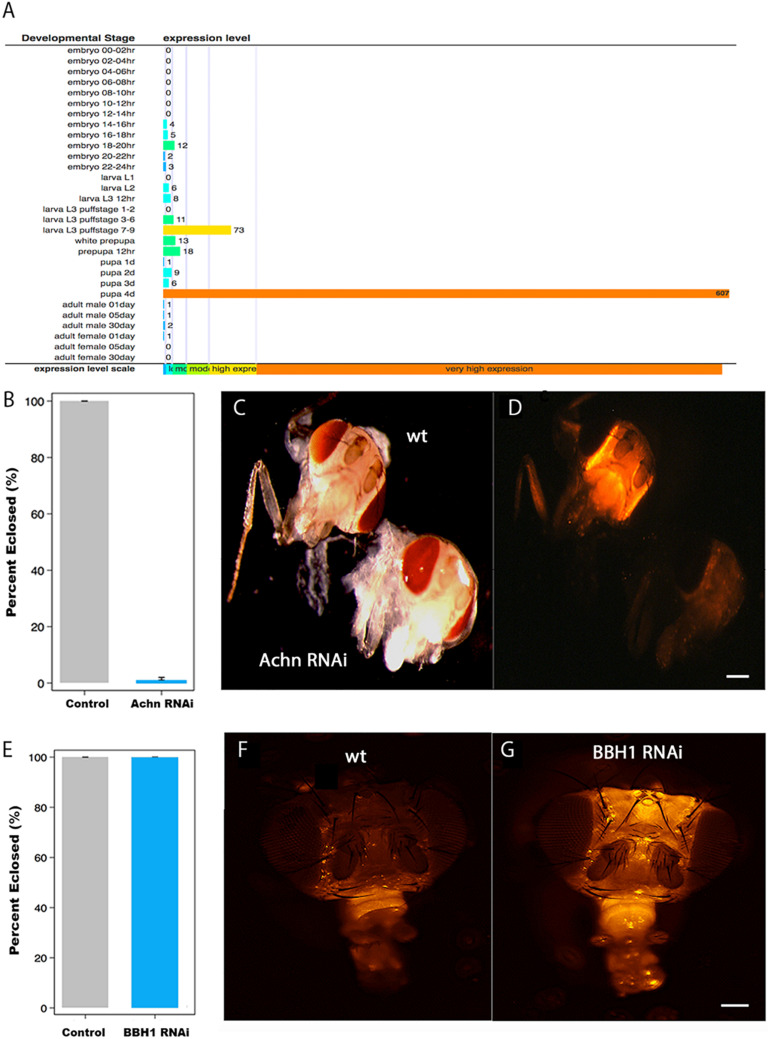
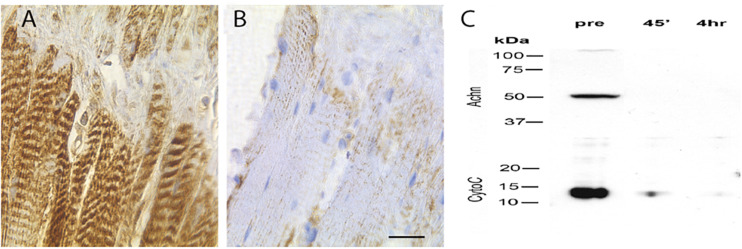
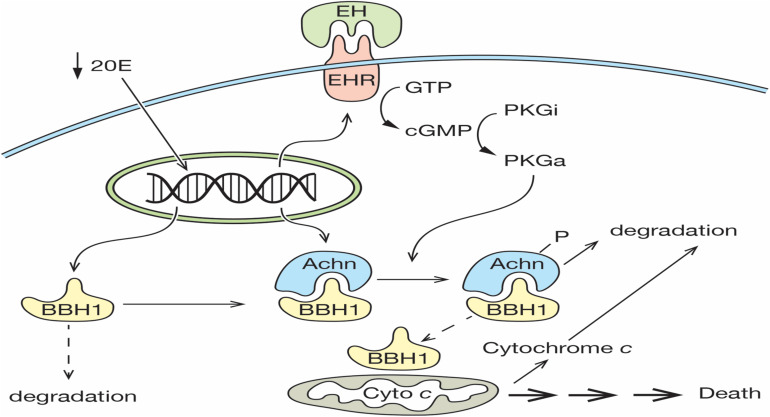
The term programmed cell death (PCD) was coined in 1965 to describe the loss of the intersegmental muscles (ISMs) of moths at the end of metamorphosis. While it was subsequently demonstrated that this hormonally controlled death requires de novo gene expression, the signal transduction pathway that couples hormone action to cell death is largely unknown. Using the ISMs from the tobacco hawkmoth Manduca sexta, we have found that Acheron/LARP6 mRNA is induced ∼1,000-fold on the day the muscles become committed to die. Acheron functions as a survival protein that protects cells until cell death is initiated at eclosion (emergence), at which point it becomes phosphorylated and degraded in response to the peptide Eclosion Hormone (EH). Acheron binds to a novel BH3-only protein that we have named BBH1 (BAD/BNIP3 homology 1). BBH1 accumulates on the day the ISMs become committed to die and is presumably liberated when Acheron is degraded. This is correlated with the release and rapid degradation of cytochrome c and the subsequent demise of the cell. RNAi experiments in the fruit fly Drosophila confirmed that loss of Acheron results in precocious ecdysial muscle death while targeting BBH1 prevents death altogether. Acheron is highly expressed in neurons and muscles in humans and drives metastatic processes in some cancers, suggesting that it may represent a novel survival protein that protects terminally differentiated cells and some cancers from death.
Keywords: BAD; BBH1; Drosophila; Manduca sexta; apoptosis; autophagy; cytochrome c; intersegmental muscle.
Copyright © 2020 Sheel, Shao, Brown, Johnson, Hamilton, Sun, Oppenheimer, Smith, Visconti, Markstein, Bigelow and Schwartz.
Figures

Similar articles
-
Differential control of cell death and gene expression during two distinct phases of hormonally-regulated muscle death in the tobacco hawkmoth Manduca sexta.J Insect Physiol. 2009 Apr;55(4):314-20. doi: 10.1016/j.jinsphys.2008.12.006. Epub 2009 Jan 29. J Insect Physiol. 2009. PMID: 19135059
-
A member of the phylogenetically conserved CAD family of transcriptional regulators is dramatically up-regulated during the programmed cell death of skeletal muscle in the tobacco hawkmoth Manduca sexta.Dev Biol. 1996 Feb 1;173(2):499-509. doi: 10.1006/dbio.1996.0043. Dev Biol. 1996. PMID: 8606008
-
Selective repression of actin and myosin heavy chain expression during the programmed death of insect skeletal muscle.Dev Biol. 1993 Aug;158(2):448-55. doi: 10.1006/dbio.1993.1202. Dev Biol. 1993. PMID: 8344462
-
Insect muscle as a model for programmed cell death.J Neurobiol. 1992 Nov;23(9):1312-26. doi: 10.1002/neu.480230918. J Neurobiol. 1992. PMID: 1469390 Review.
-
Thinking globally, acting locally: steroid hormone regulation of the dendritic architecture, synaptic connectivity and death of an individual neuron.Prog Neurobiol. 2003 Aug;70(5):421-42. doi: 10.1016/s0301-0082(03)00102-3. Prog Neurobiol. 2003. PMID: 14511700 Review.
Cited by
-
LARP6 suppresses colorectal cancer progression through ZNF267/SGMS2-mediated imbalance of sphingomyelin synthesis.J Exp Clin Cancer Res. 2023 Jan 24;42(1):33. doi: 10.1186/s13046-023-02605-4. J Exp Clin Cancer Res. 2023. PMID: 36691044 Free PMC article.
-
High-resolution analysis of differential gene expression during skeletal muscle atrophy and programmed cell death.Physiol Genomics. 2020 Oct 1;52(10):492-511. doi: 10.1152/physiolgenomics.00047.2020. Epub 2020 Sep 14. Physiol Genomics. 2020. PMID: 32926651 Free PMC article.
-
Somatic piRNAs and Transposons are Differentially Expressed Coincident with Skeletal Muscle Atrophy and Programmed Cell Death.Front Genet. 2021 Dec 22;12:775369. doi: 10.3389/fgene.2021.775369. eCollection 2021. Front Genet. 2021. PMID: 35003216 Free PMC article.
-
SFPQ and Its Isoform as Potential Biomarker for Non-Small-Cell Lung Cancer.Int J Mol Sci. 2023 Aug 6;24(15):12500. doi: 10.3390/ijms241512500. Int J Mol Sci. 2023. PMID: 37569873 Free PMC article.
-
Genome-Wide Gene Expression Profiles Reveal Distinct Molecular Characteristics of the Goose Granulosa Cells.Front Genet. 2021 Dec 17;12:786287. doi: 10.3389/fgene.2021.786287. eCollection 2021. Front Genet. 2021. PMID: 34992633 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials