

Silencing and Transcriptional Regulation of Endogenous Retroviruses: An Overview
- PMID: 32823517
- PMCID: PMC7472088
- DOI: 10.3390/v12080884
Silencing and Transcriptional Regulation of Endogenous Retroviruses: An Overview
Abstract
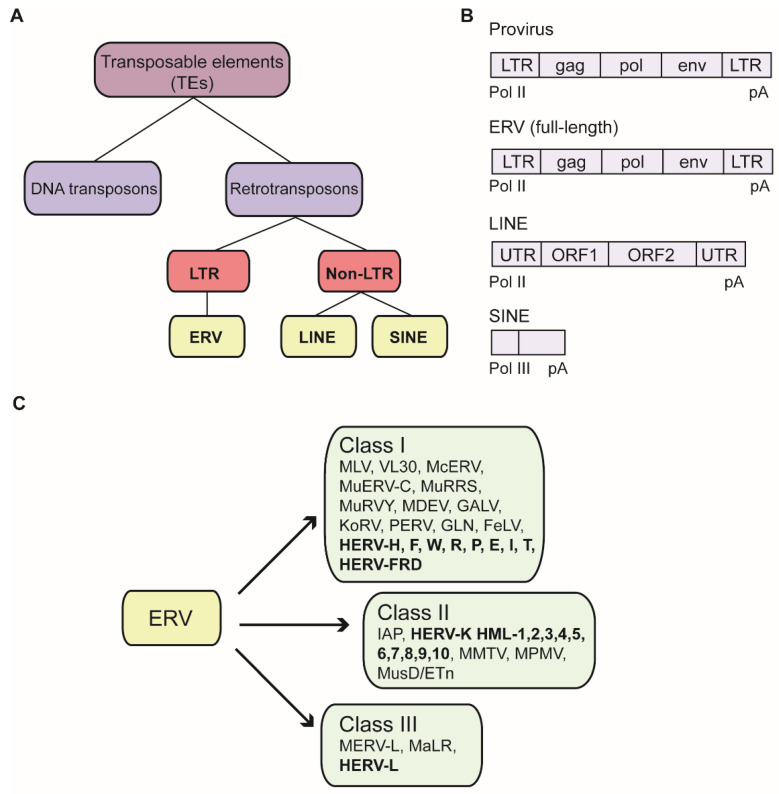
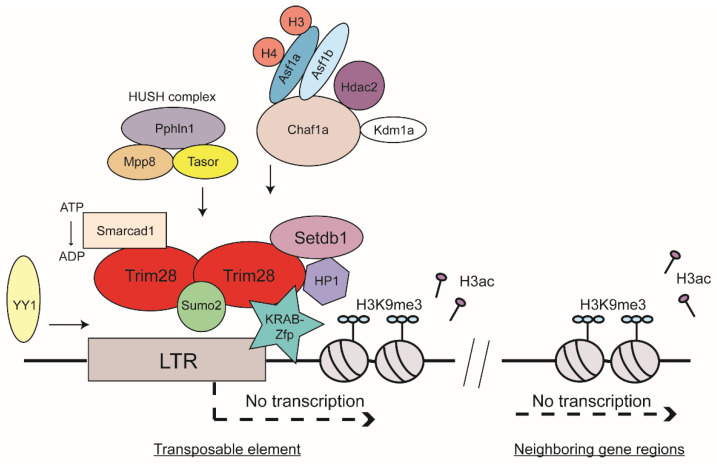
Almost half of the human genome is made up of transposable elements (TEs), and about 8% consists of endogenous retroviruses (ERVs). ERVs are remnants of ancient exogenous retrovirus infections of the germ line. Most TEs are inactive and not detrimental to the host. They are tightly regulated to ensure genomic stability of the host and avoid deregulation of nearby gene loci. Histone-based posttranslational modifications such as H3K9 trimethylation are one of the main silencing mechanisms. Trim28 is one of the identified master regulators of silencing, which recruits most prominently the H3K9 methyltransferase Setdb1, among other factors. Sumoylation and ATP-dependent chromatin remodeling factors seem to contribute to proper localization of Trim28 to ERV sequences and promote Trim28 interaction with Setdb1. Additionally, DNA methylation as well as RNA-mediated targeting of TEs such as piRNA-based silencing play important roles in ERV regulation. Despite the involvement of ERV overexpression in several cancer types, autoimmune diseases, and viral pathologies, ERVs are now also appreciated for their potential positive role in evolution. ERVs can provide new regulatory gene elements or novel binding sites for transcription factors, and ERV gene products can even be repurposed for the benefit of the host.
Keywords: ERV expression and diseases; co-option of ERV functions; endogenous retroviruses (ERV); transcriptional silencing; transposable elements.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Silencing of endogenous retroviruses by heterochromatin.Cell Mol Life Sci. 2017 Jun;74(11):2055-2065. doi: 10.1007/s00018-017-2454-8. Epub 2017 Feb 3. Cell Mol Life Sci. 2017. PMID: 28160052 Free PMC article. Review.
-
hnRNP K coordinates transcriptional silencing by SETDB1 in embryonic stem cells.PLoS Genet. 2015 Jan 22;11(1):e1004933. doi: 10.1371/journal.pgen.1004933. eCollection 2015 Jan. PLoS Genet. 2015. PMID: 25611934 Free PMC article.
-
Existence of Two Distinct Infectious Endogenous Retroviruses in Domestic Cats and Their Different Strategies for Adaptation to Transcriptional Regulation.J Virol. 2016 Sep 29;90(20):9029-45. doi: 10.1128/JVI.00716-16. Print 2016 Oct 15. J Virol. 2016. PMID: 27466428 Free PMC article.
-
The histone methyltransferase SETDB1 represses endogenous and exogenous retroviruses in B lymphocytes.Proc Natl Acad Sci U S A. 2015 Jul 7;112(27):8367-72. doi: 10.1073/pnas.1422187112. Epub 2015 Jun 22. Proc Natl Acad Sci U S A. 2015. PMID: 26100872 Free PMC article.
-
Endogenous Retroviruses Walk a Fine Line between Priming and Silencing.Viruses. 2020 Jul 23;12(8):792. doi: 10.3390/v12080792. Viruses. 2020. PMID: 32718022 Free PMC article. Review.
Cited by
-
The role of human endogenous retroviruses in gliomas: from etiological perspectives and therapeutic implications.Neuro Oncol. 2021 Oct 1;23(10):1647-1655. doi: 10.1093/neuonc/noab142. Neuro Oncol. 2021. PMID: 34120190 Free PMC article. Review.
-
Unveiling the molecular structure and role of RBBP4/7: implications for epigenetic regulation and cancer research.Front Mol Biosci. 2023 Nov 13;10:1276612. doi: 10.3389/fmolb.2023.1276612. eCollection 2023. Front Mol Biosci. 2023. PMID: 38028543 Free PMC article. Review.
-
HERV-K Envelope Protein Induces Long-Lasting Production of Autoantibodies in T1DM Patients at Onset in Comparison to ZNT8 Autoantibodies.Pathogens. 2022 Oct 15;11(10):1188. doi: 10.3390/pathogens11101188. Pathogens. 2022. PMID: 36297245 Free PMC article.
-
Transcriptional and reverse transcriptional regulation of host genes by human endogenous retroviruses in cancers.Front Microbiol. 2022 Jul 19;13:946296. doi: 10.3389/fmicb.2022.946296. eCollection 2022. Front Microbiol. 2022. PMID: 35928153 Free PMC article. Review.
-
Insights into Mechanisms of Tumorigenesis in Neuroendocrine Neoplasms.Int J Mol Sci. 2021 Sep 25;22(19):10328. doi: 10.3390/ijms221910328. Int J Mol Sci. 2021. PMID: 34638668 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
