

IκBα Nuclear Export Enables 4-1BB-Induced cRel Activation and IL-2 Production to Promote CD8 T Cell Immunity
- PMID: 32817348
- PMCID: PMC7484350
- DOI: 10.4049/jimmunol.2000039
IκBα Nuclear Export Enables 4-1BB-Induced cRel Activation and IL-2 Production to Promote CD8 T Cell Immunity
Abstract
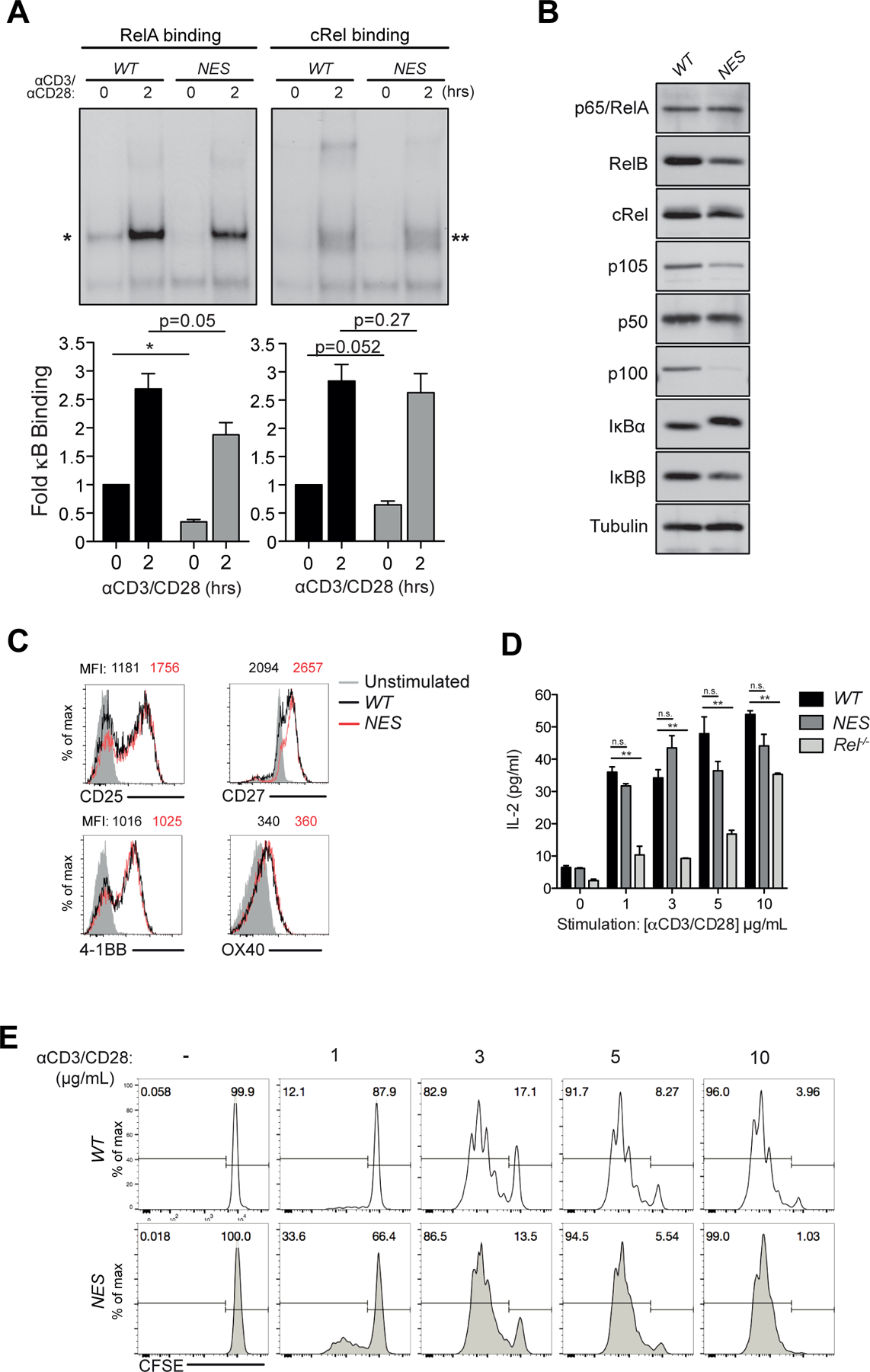
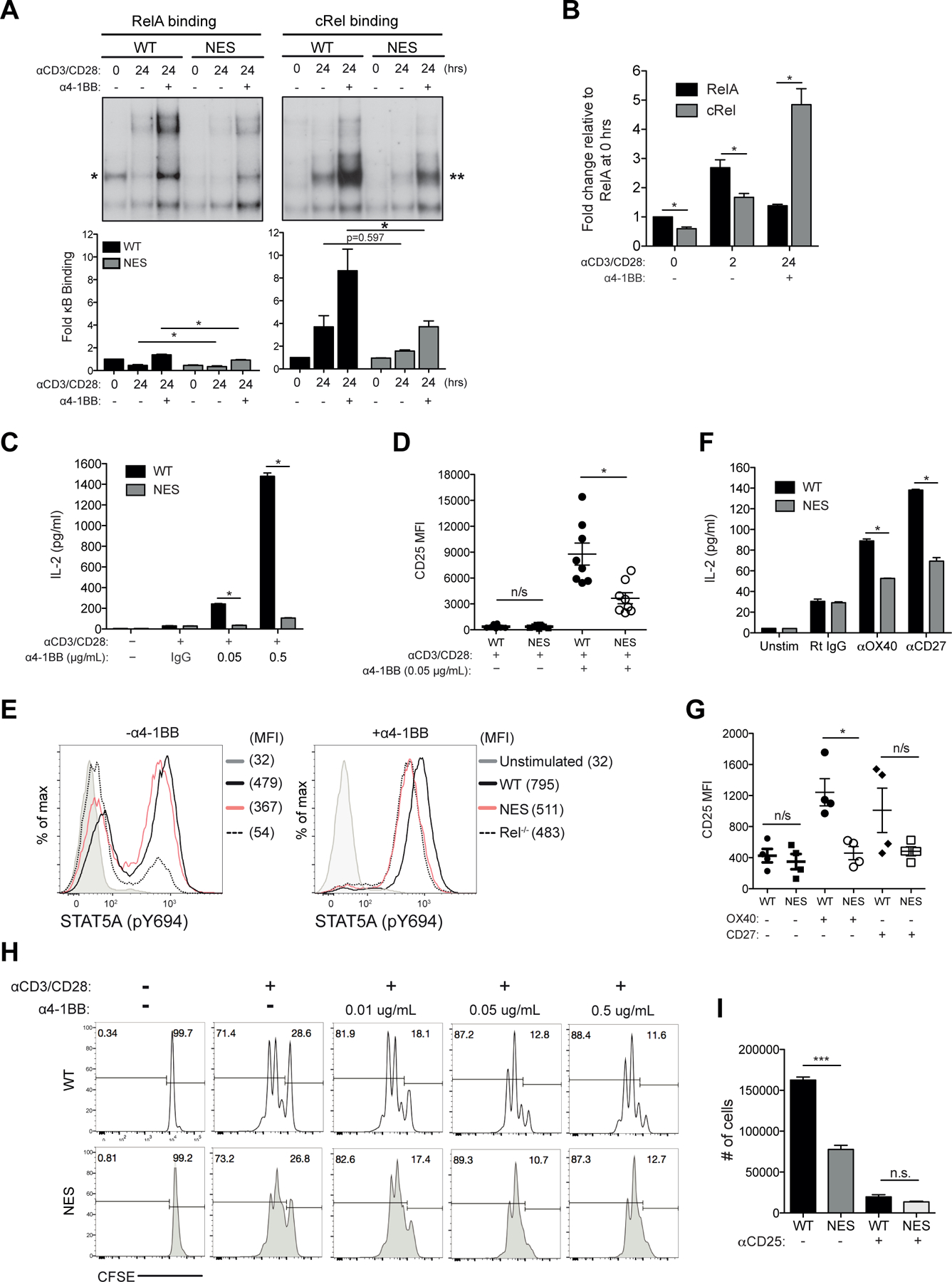
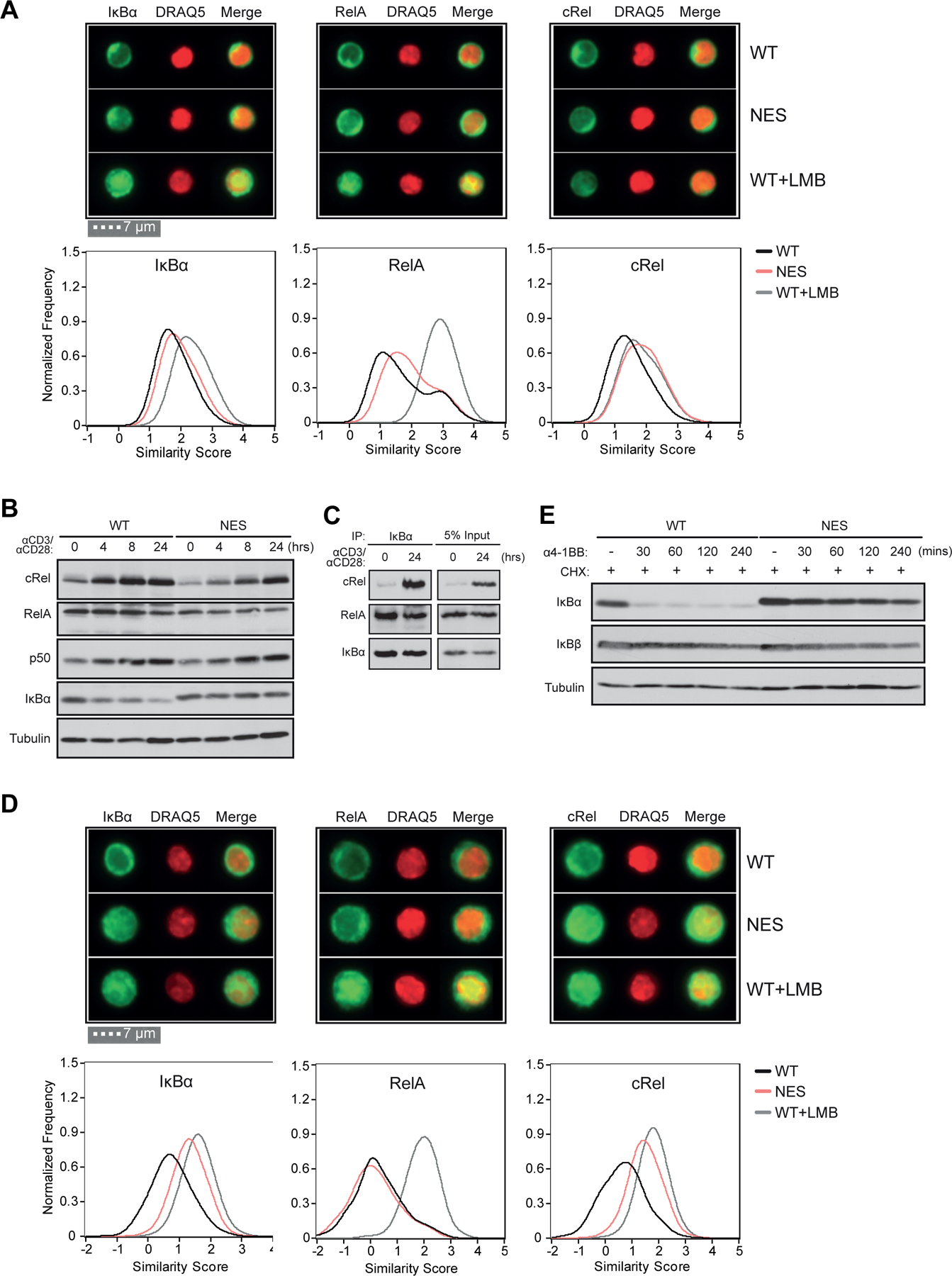
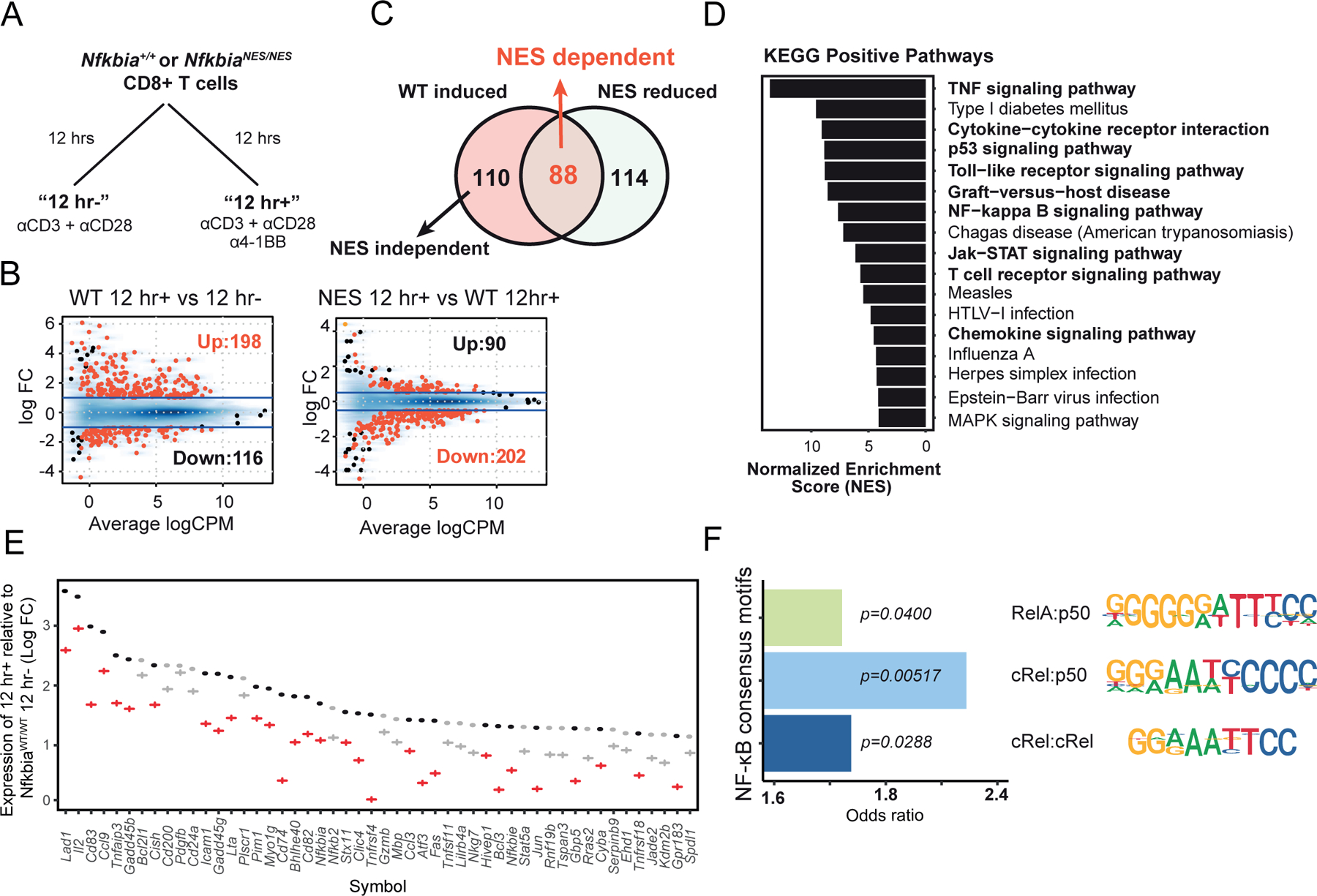
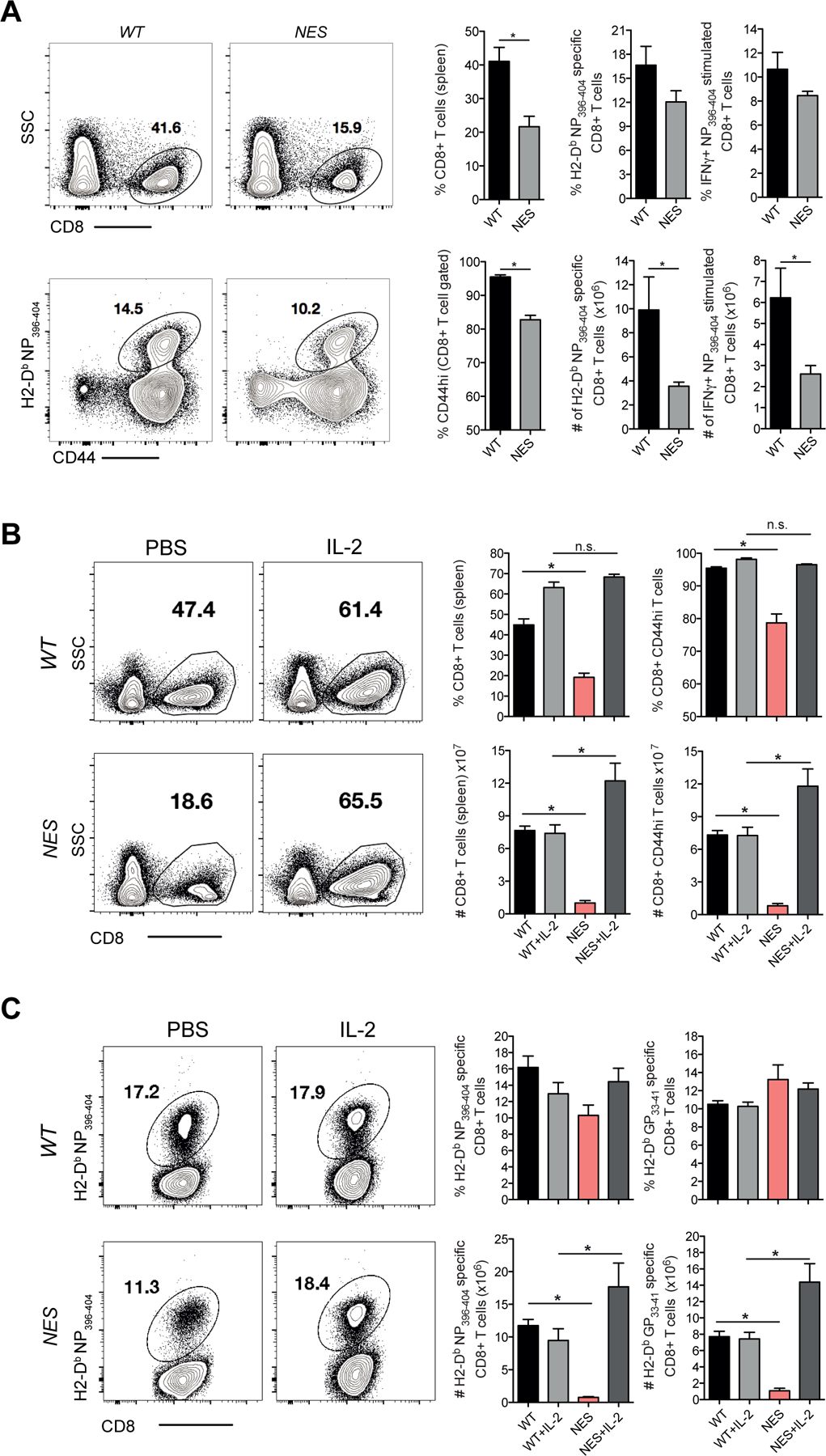
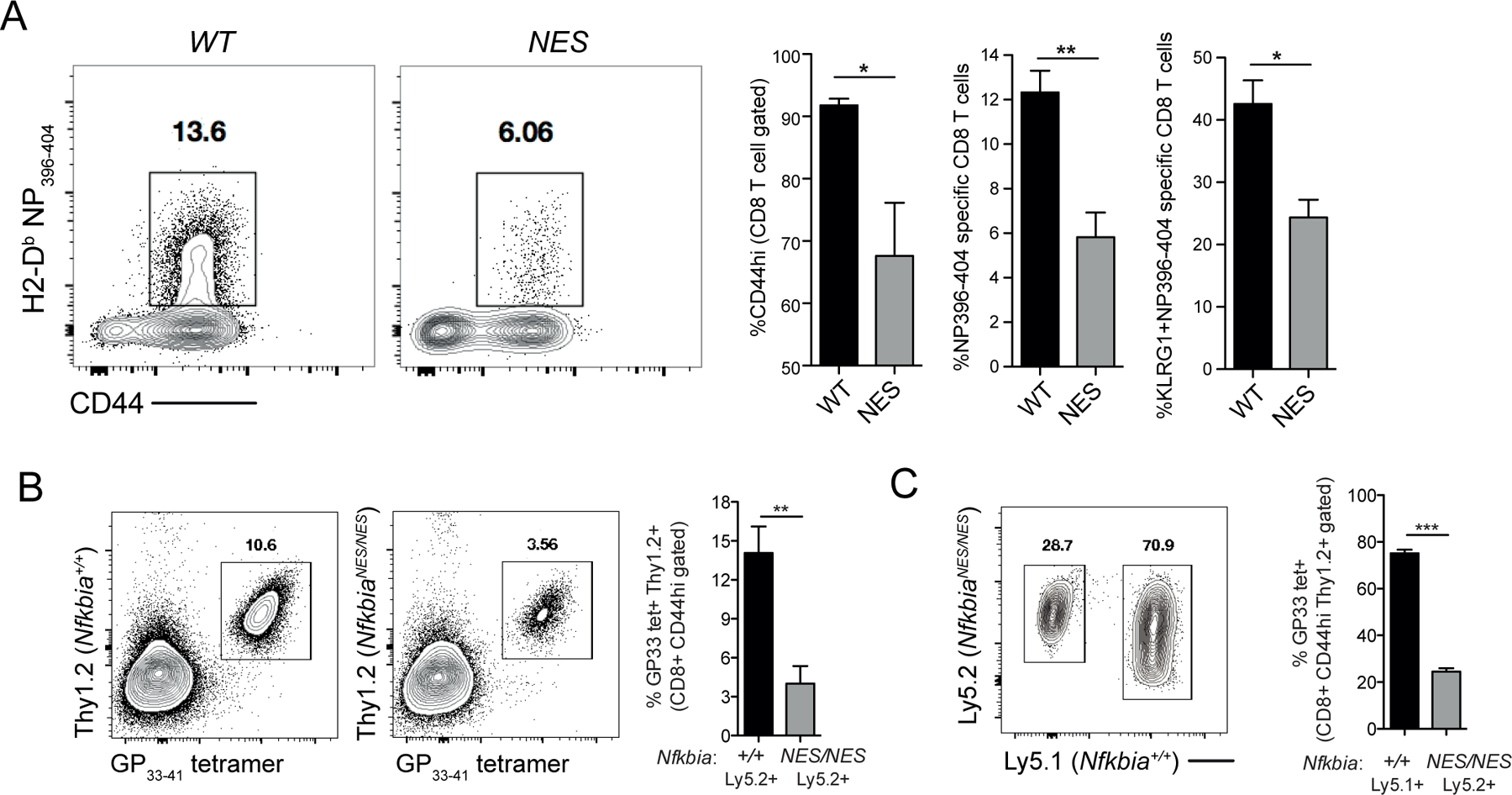
Optimal CD8 T cell immunity is orchestrated by signaling events initiated by TCR recognition of peptide Ag in concert with signals from molecules such as CD28 and 4-1BB. The molecular mechanisms underlying the temporal and spatial signaling dynamics in CD8 T cells remain incompletely understood. In this study, we show that stimulation of naive CD8 T cells with agonistic CD3 and CD28 Abs, mimicking TCR and costimulatory signals, coordinately induces 4-1BB and cRel to enable elevated cytosolic cRel:IκBα complex formation and subsequent 4-1BB-induced IκBα degradation, sustained cRel activation, heightened IL-2 production and T cell expansion. NfkbiaNES/NES CD8 T cells harboring a mutated IκBα nuclear export sequence abnormally accumulate inactive cRel:IκBα complexes in the nucleus following stimulation with agonistic anti-CD3 and anti-CD28 Abs, rendering them resistant to 4-1BB induced signaling and a disrupted chain of events necessary for efficient T cell expansion. Consequently, CD8 T cells in NfkbiaNES/NES mice poorly expand during viral infection, and this can be overcome by exogenous IL-2 administration. Consistent with cell-based data, adoptive transfer experiments demonstrated that the antiviral CD8 T cell defect in NfkbiaNES/NES mice was cell intrinsic. Thus, these results reveal that IκBα, via its unique nuclear export function, enables, rather than inhibits 4-1BB-induced cRel activation and IL-2 production to facilitate optimal CD8 T cell immunity.
Copyright © 2020 by The American Association of Immunologists, Inc.
Conflict of interest statement
Conflicts of Interest
The authors declare no competing financial interests.
Figures
Similar articles
-
4-1BB ligand induces cell division, sustains survival, and enhances effector function of CD4 and CD8 T cells with similar efficacy.J Immunol. 2001 Aug 1;167(3):1313-24. doi: 10.4049/jimmunol.167.3.1313. J Immunol. 2001. PMID: 11466348
-
Mechanism responsible for T-cell antigen receptor- and CD28- or interleukin 1 (IL-1) receptor-initiated regulation of IL-2 gene expression by NF-kappaB.Mol Cell Biol. 1998 Jun;18(6):3140-8. doi: 10.1128/MCB.18.6.3140. Mol Cell Biol. 1998. PMID: 9584155 Free PMC article.
-
Co-stimulation through 4-1BB/CD137 improves the expansion and function of CD8(+) melanoma tumor-infiltrating lymphocytes for adoptive T-cell therapy.PLoS One. 2013;8(4):e60031. doi: 10.1371/journal.pone.0060031. Epub 2013 Apr 1. PLoS One. 2013. PMID: 23560068 Free PMC article.
-
Costimulation through the CD137/4-1BB pathway protects human melanoma tumor-infiltrating lymphocytes from activation-induced cell death and enhances antitumor effector function.J Immunother. 2011 Apr;34(3):236-50. doi: 10.1097/CJI.0b013e318209e7ec. J Immunother. 2011. PMID: 21389874 Free PMC article.
-
4-1BB is superior to CD28 costimulation for generating CD8+ cytotoxic lymphocytes for adoptive immunotherapy.J Immunol. 2007 Oct 1;179(7):4910-8. doi: 10.4049/jimmunol.179.7.4910. J Immunol. 2007. PMID: 17878391 Free PMC article.
Cited by
-
TFAP2A-activated ITGB4 promotes lung adenocarcinoma progression and inhibits CD4+/CD8+ T-cell infiltrations by targeting NF-κB signaling pathway.Transl Lung Cancer Res. 2024 Sep 30;13(9):2116-2138. doi: 10.21037/tlcr-24-50. Epub 2024 Sep 10. Transl Lung Cancer Res. 2024. PMID: 39430326 Free PMC article.
-
Uncovering receptor-ligand interactions using a high-avidity CRISPR activation screening platform.Sci Adv. 2024 Feb 16;10(7):eadj2445. doi: 10.1126/sciadv.adj2445. Epub 2024 Feb 14. Sci Adv. 2024. PMID: 38354234 Free PMC article.
-
An OX40L mRNA vaccine inhibits the growth of hepatocellular carcinoma.Front Oncol. 2022 Oct 13;12:975408. doi: 10.3389/fonc.2022.975408. eCollection 2022. Front Oncol. 2022. PMID: 36313716 Free PMC article.
-
A gain-of-function p53 mutant synergizes with oncogenic NRAS to promote acute myeloid leukemia in mice.J Clin Invest. 2023 Dec 15;133(24):e173116. doi: 10.1172/JCI173116. J Clin Invest. 2023. PMID: 37847561 Free PMC article.
-
Valosin-containing protein (VCP/p97) inhibition reduces viral clearance and induces toxicity associated with muscular damage.Cell Death Dis. 2022 Dec 1;13(12):1015. doi: 10.1038/s41419-022-05461-w. Cell Death Dis. 2022. PMID: 36456548 Free PMC article.
References
-
- Borowski AB, Boesteanu AC, Mueller YM, Carafides C, Topham DJ, Altman JD, Jennings SR, and Katsikis PD. 2007. Memory CD8+ T cells require CD28 costimulation. J Immunol 179: 6494–6503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials