

Centromeres under Pressure: Evolutionary Innovation in Conflict with Conserved Function
- PMID: 32784998
- PMCID: PMC7463522
- DOI: 10.3390/genes11080912
Centromeres under Pressure: Evolutionary Innovation in Conflict with Conserved Function
Abstract
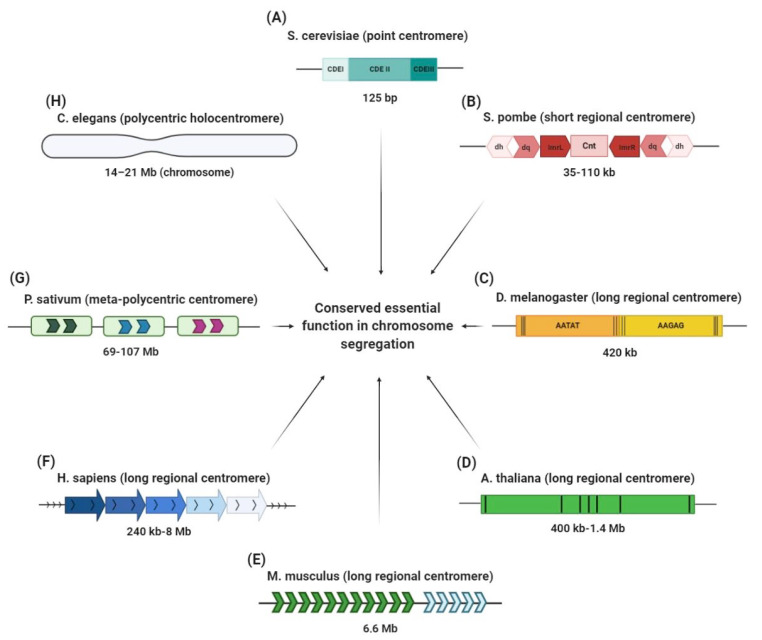
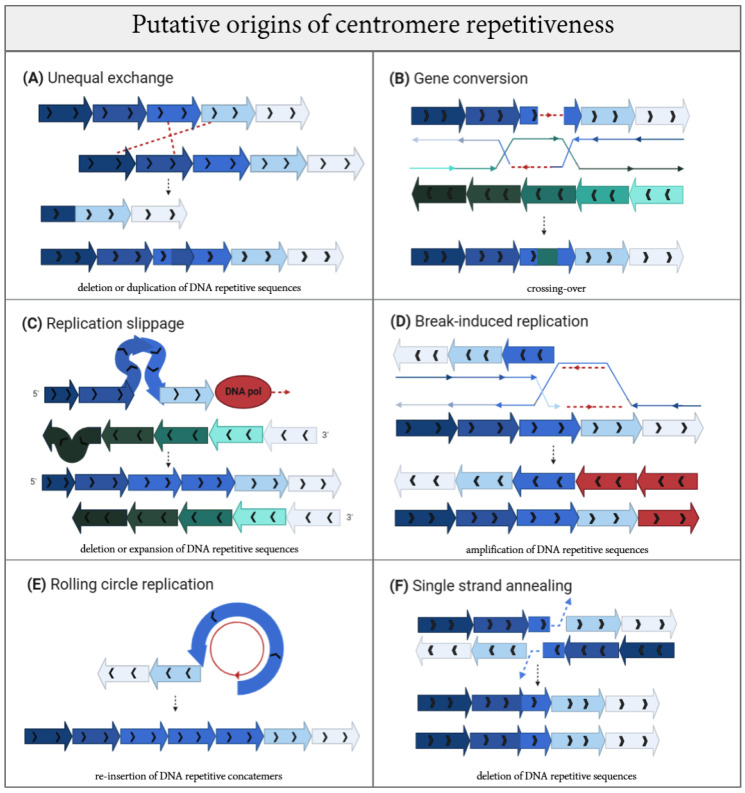
Centromeres are essential genetic elements that enable spindle microtubule attachment for chromosome segregation during mitosis and meiosis. While this function is preserved across species, centromeres display an array of dynamic features, including: (1) rapidly evolving DNA; (2) wide evolutionary diversity in size, shape and organization; (3) evidence of mutational processes to generate homogenized repetitive arrays that characterize centromeres in several species; (4) tolerance to changes in position, as in the case of neocentromeres; and (5) intrinsic fragility derived by sequence composition and secondary DNA structures. Centromere drive underlies rapid centromere DNA evolution due to the "selfish" pursuit to bias meiotic transmission and promote the propagation of stronger centromeres. Yet, the origins of other dynamic features of centromeres remain unclear. Here, we review our current understanding of centromere evolution and plasticity. We also detail the mutagenic processes proposed to shape the divergent genetic nature of centromeres. Changes to centromeres are not simply evolutionary relics, but ongoing shifts that on one side promote centromere flexibility, but on the other can undermine centromere integrity and function with potential pathological implications such as genome instability.
Keywords: HORs; centromere; centromere evolution; chromosome instability; mutagenesis; repetitive DNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Repetitive Elements Contribute to the Diversity and Evolution of Centromeres in the Fungal Genus Verticillium.mBio. 2020 Sep 8;11(5):e01714-20. doi: 10.1128/mBio.01714-20. mBio. 2020. PMID: 32900804 Free PMC article.
-
Centromeres Drive a Hard Bargain.Trends Genet. 2017 Feb;33(2):101-117. doi: 10.1016/j.tig.2016.12.001. Epub 2017 Jan 7. Trends Genet. 2017. PMID: 28069312 Free PMC article. Review.
-
Molecular Strategies of Meiotic Cheating by Selfish Centromeres.Cell. 2019 Aug 22;178(5):1132-1144.e10. doi: 10.1016/j.cell.2019.07.001. Epub 2019 Aug 8. Cell. 2019. PMID: 31402175 Free PMC article.
-
The birth of the centromere.Cell Cycle. 2007 Dec 1;6(23):2872-6. doi: 10.4161/cc.6.23.5047. Cell Cycle. 2007. PMID: 18156801 Review.
-
Epigenetics as an Evolutionary Tool for Centromere Flexibility.Genes (Basel). 2020 Jul 16;11(7):809. doi: 10.3390/genes11070809. Genes (Basel). 2020. PMID: 32708654 Free PMC article. Review.
Cited by
-
Emerging Single-Cell Technological Approaches to Investigate Chromatin Dynamics and Centromere Regulation in Human Health and Disease.Int J Mol Sci. 2021 Aug 16;22(16):8809. doi: 10.3390/ijms22168809. Int J Mol Sci. 2021. PMID: 34445507 Free PMC article. Review.
-
Cytogenetic Analysis of Satellitome of Madagascar Leaf-Tailed Geckos.Genes (Basel). 2024 Mar 28;15(4):429. doi: 10.3390/genes15040429. Genes (Basel). 2024. PMID: 38674364 Free PMC article.
-
Karyotype and LTR-RTs analysis provide insights into oak genomic evolution.BMC Genomics. 2024 Apr 3;25(1):328. doi: 10.1186/s12864-024-10177-6. BMC Genomics. 2024. PMID: 38566015 Free PMC article.
-
The complete diploid reference genome of RPE-1 identifies human phased epigenetic landscapes.bioRxiv [Preprint]. 2023 Dec 30:2023.11.01.565049. doi: 10.1101/2023.11.01.565049. bioRxiv. 2023. PMID: 38168337 Free PMC article. Preprint.
-
Molecular mechanisms of aging and anti-aging strategies.Cell Commun Signal. 2024 May 24;22(1):285. doi: 10.1186/s12964-024-01663-1. Cell Commun Signal. 2024. PMID: 38790068 Free PMC article. Review.
References
-
- Flemming W. Zellsubstanz, Kern und Zelltheilung. DMW—Dtsch. Med. Wochenschr. 1883;9:342. doi: 10.1055/s-0029-1197237. - DOI
-
- Darlington C.D., Hallpike C.S., Hartridge H., Rawdon-Smith A.F. The external mechanics of the chromosomes I—The scope of enquiry. Proc. R. Soc. Lond. Ser. B: Biol. Sci. 1936;121:264–273. doi: 10.1098/rspb.1936.0064. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
