

Tolerogenic vaccines: Targeting the antigenic and cytokine niches of FOXP3+ regulatory T cells
- PMID: 32712270
- PMCID: PMC7444458
- DOI: 10.1016/j.cellimm.2020.104173
Tolerogenic vaccines: Targeting the antigenic and cytokine niches of FOXP3+ regulatory T cells
Abstract
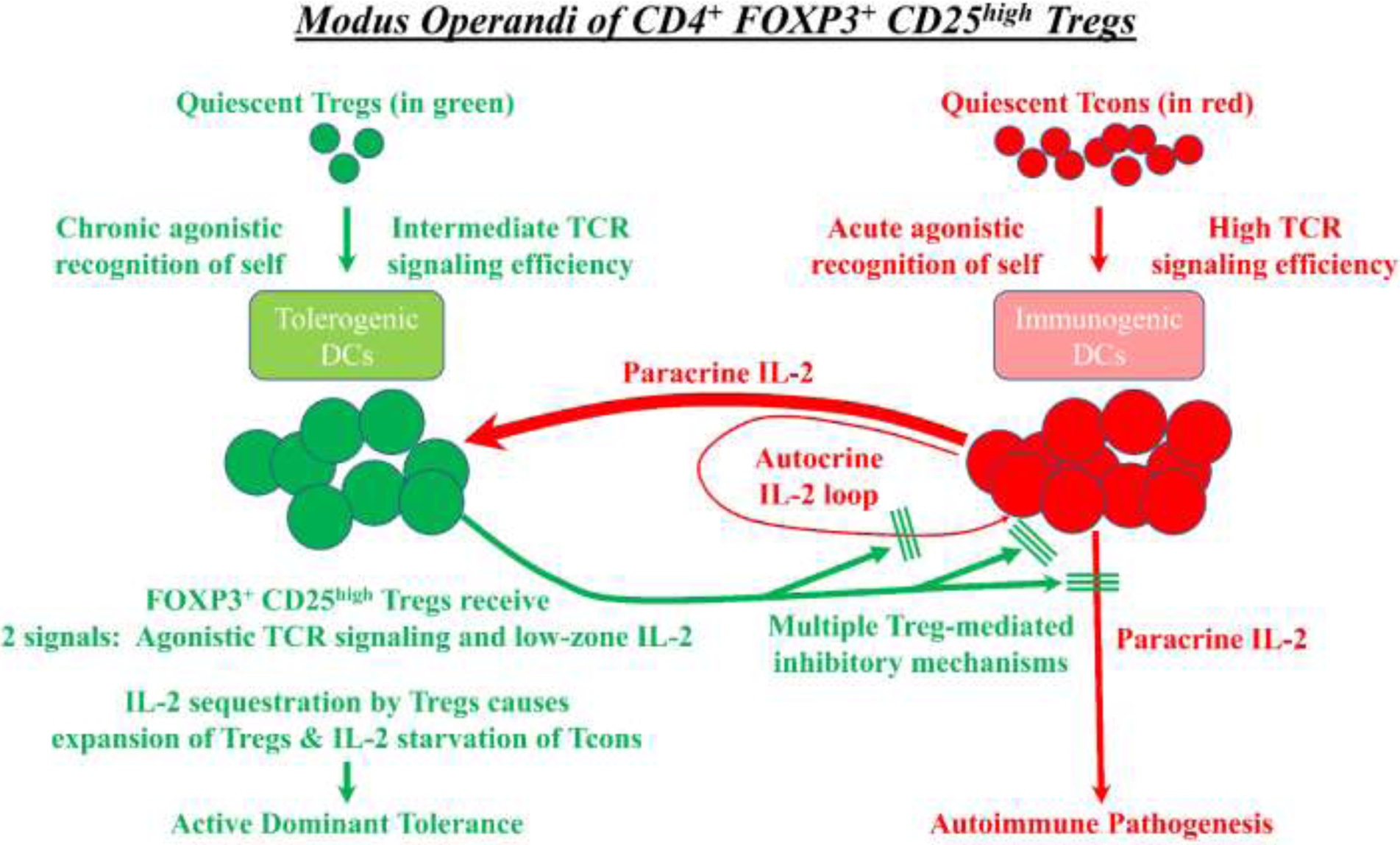
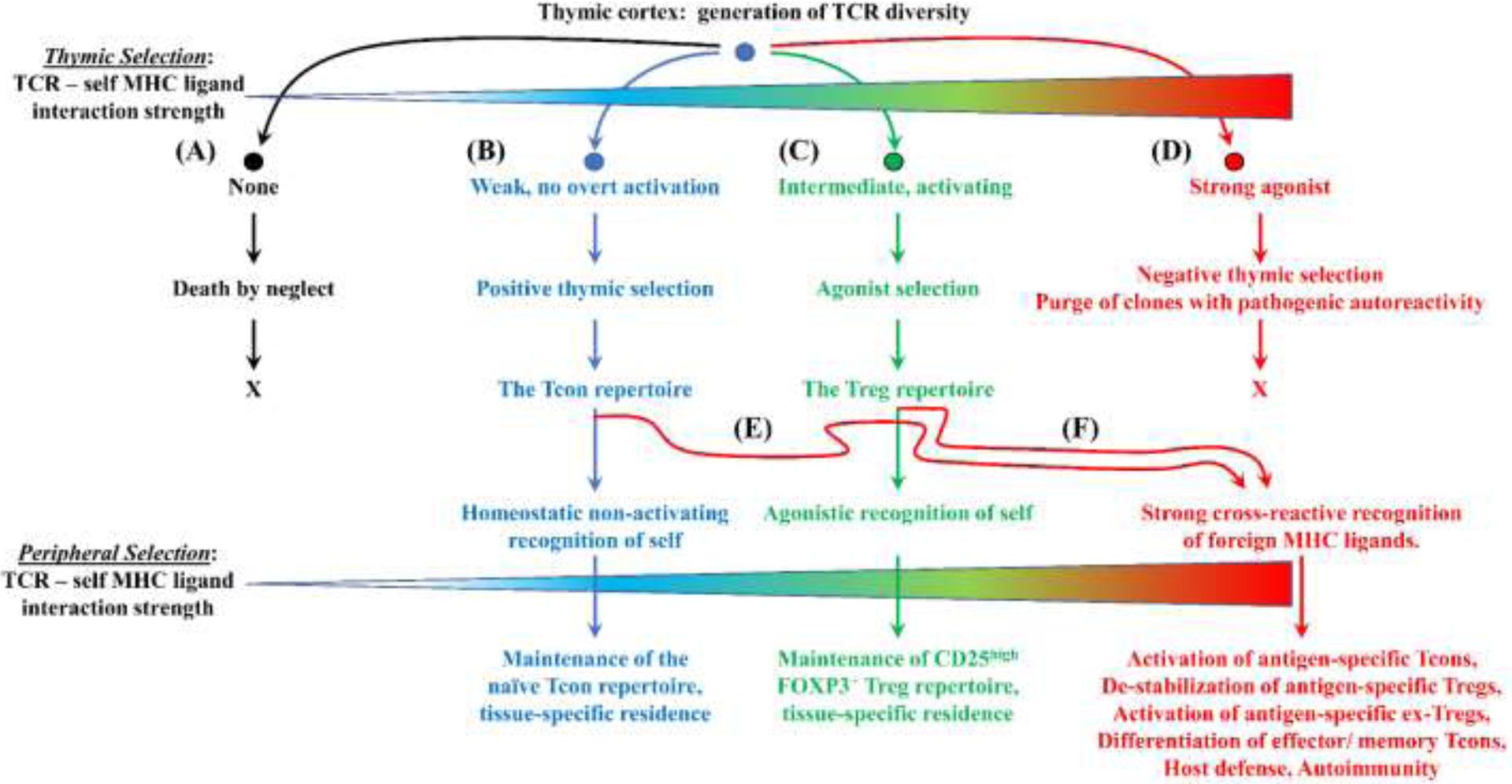
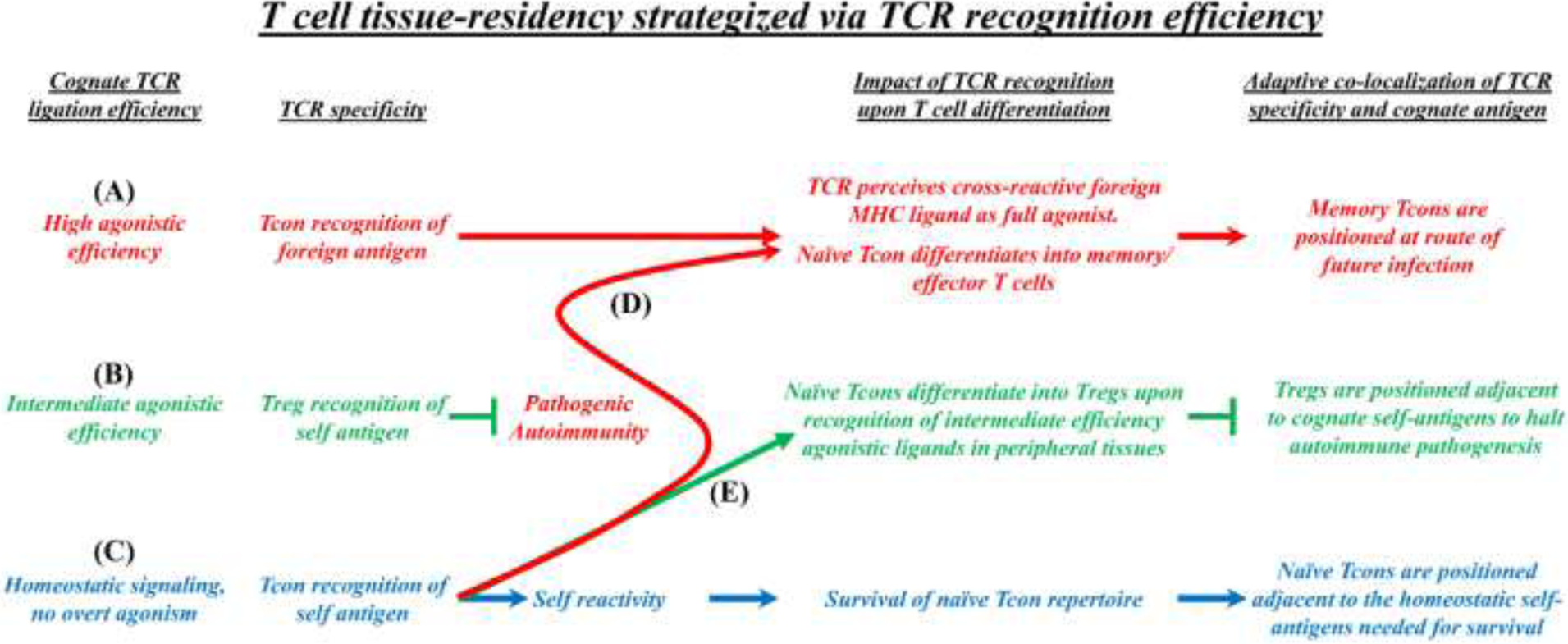
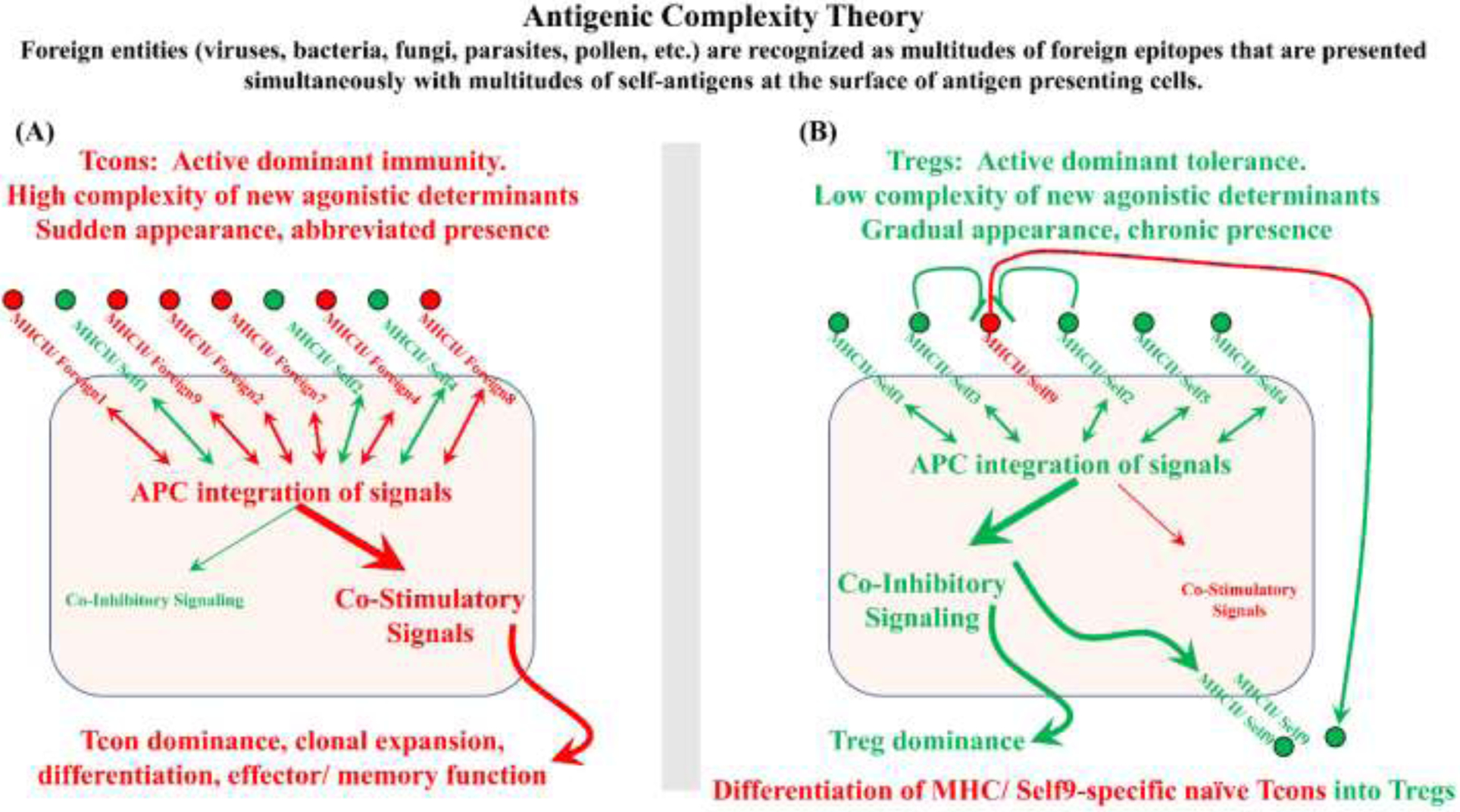
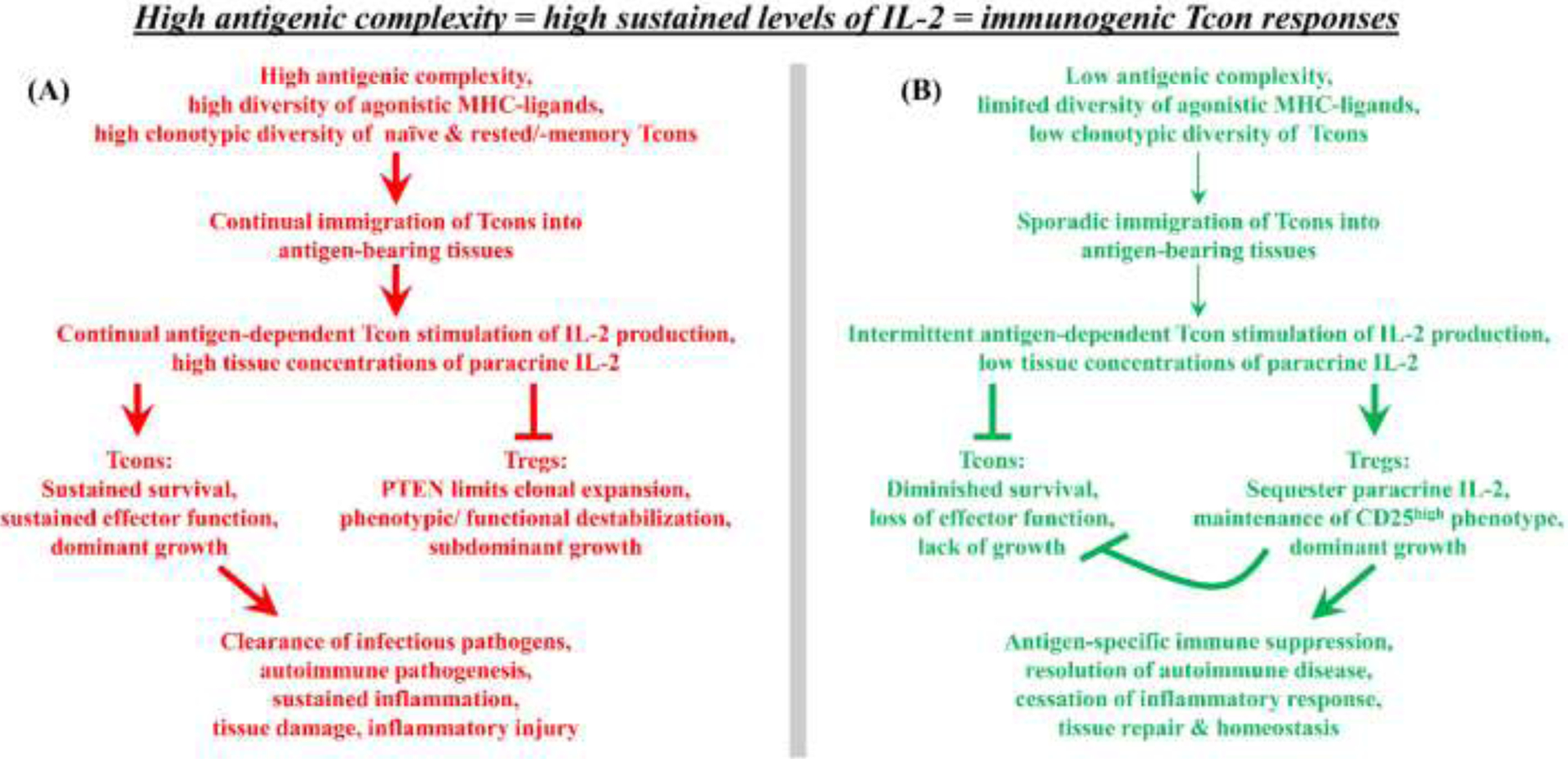
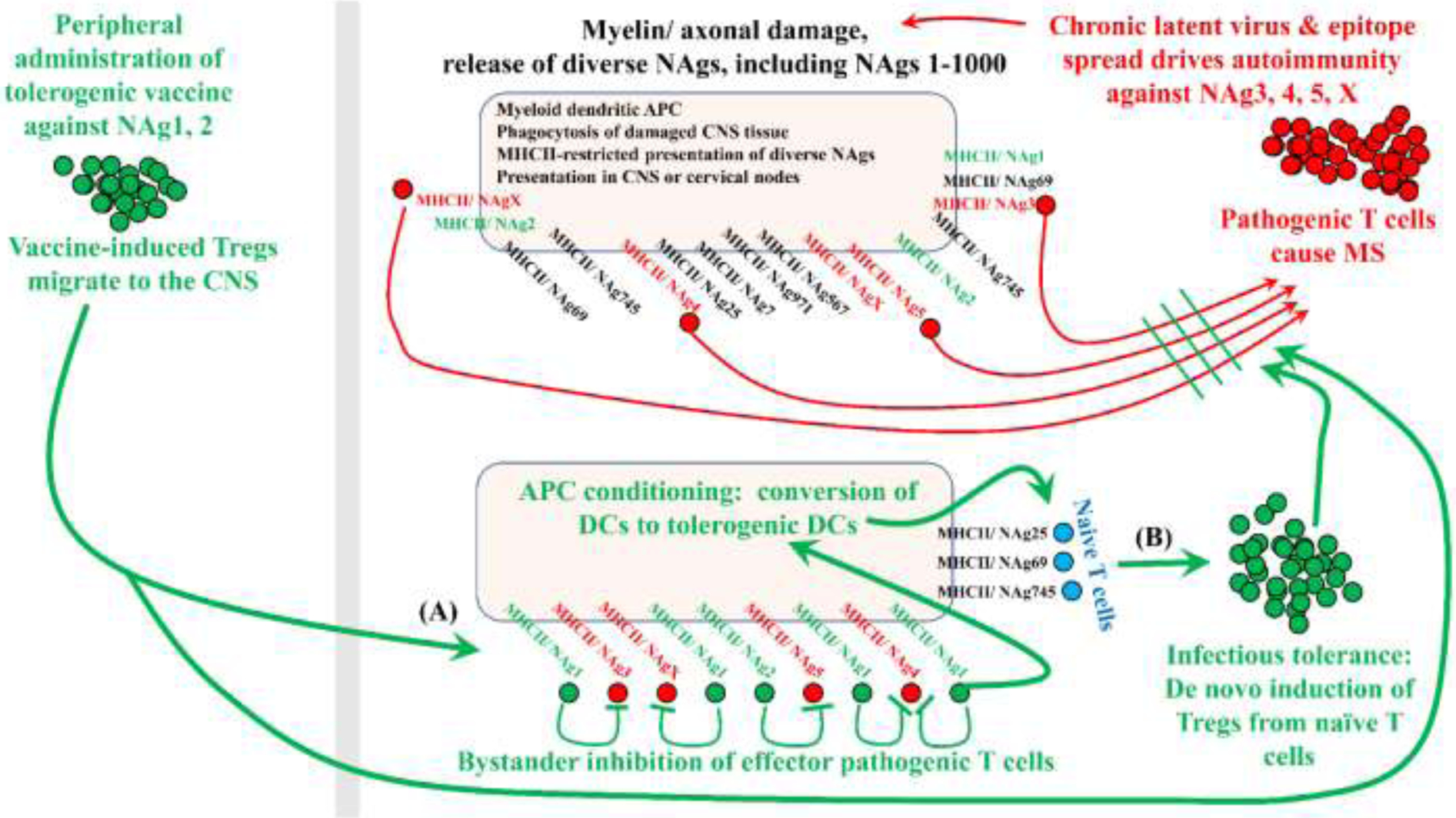
FOXP3+ regulatory T cells (Tregs) constitute a critical barrier that enforces tolerance to both the self-peptidome and the extended-self peptidome to ensure tissue-specific resistance to autoimmune, allergic, and other inflammatory disorders. Here, we review intuitive models regarding how T cell antigen receptor (TCR) specificity and antigen recognition efficiency shape the Treg and conventional T cell (Tcon) repertoires to adaptively regulate T cell maintenance, tissue-residency, phenotypic stability, and immune function in peripheral tissues. Three zones of TCR recognition efficiency are considered, including Tcon recognition of specific low-efficiency self MHC-ligands, Treg recognition of intermediate-efficiency agonistic self MHC-ligands, and Tcon recognition of cross-reactive high-efficiency agonistic foreign MHC-ligands. These respective zones of TCR recognition efficiency are key to understanding how tissue-resident immune networks integrate the antigenic complexity of local environments to provide adaptive decisions setting the balance of suppressive and immunogenic responses. Importantly, deficiencies in the Treg repertoire appear to be an important cause of chronic inflammatory disease. Deficiencies may include global deficiencies in Treg numbers or function, subtle 'holes in the Treg repertoire' in tissue-resident Treg populations, or simply Treg insufficiencies that are unable to counter an overwhelming molecular mimicry stimulus. Tolerogenic vaccination and Treg-based immunotherapy are two therapeutic modalities meant to restore dominance of Treg networks to reverse chronic inflammatory disease. Studies of these therapeutic modalities in a preclinical setting have provided insight into the Treg niche, including the concept that intermediate-efficiency TCR signaling, high IFN-β concentrations, and low IL-2 concentrations favor Treg responses and active dominant mechanisms of immune tolerance. Overall, the purpose here is to assimilate new and established concepts regarding how cognate TCR specificity of the Treg repertoire and the contingent cytokine networks provide a foundation for understanding Treg suppressive strategy.
Keywords: Clonotypic specificity; FOXP3(+) Tregs; Immunological tolerance; T cell antigen receptor; Tolerogenic vaccination.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
A GMCSF-Neuroantigen Tolerogenic Vaccine Elicits Systemic Lymphocytosis of CD4+ CD25high FOXP3+ Regulatory T Cells in Myelin-Specific TCR Transgenic Mice Contingent Upon Low-Efficiency T Cell Antigen Receptor Recognition.Front Immunol. 2019 Jan 10;9:3119. doi: 10.3389/fimmu.2018.03119. eCollection 2018. Front Immunol. 2019. PMID: 30687323 Free PMC article.
-
A GM-CSF-neuroantigen tolerogenic vaccine elicits inefficient antigen recognition events below the CD40L triggering threshold to expand CD4+ CD25+ FOXP3+ Tregs that inhibit experimental autoimmune encephalomyelitis (EAE).J Neuroinflammation. 2020 Jun 10;17(1):180. doi: 10.1186/s12974-020-01856-8. J Neuroinflammation. 2020. PMID: 32522287 Free PMC article.
-
Thymic commitment of regulatory T cells is a pathway of TCR-dependent selection that isolates repertoires undergoing positive or negative selection.Curr Top Microbiol Immunol. 2005;293:43-71. doi: 10.1007/3-540-27702-1_3. Curr Top Microbiol Immunol. 2005. PMID: 15981475 Review.
-
Regulatory T cells: roles of T cell receptor for their development and function.Semin Immunopathol. 2010 Jun;32(2):95-106. doi: 10.1007/s00281-010-0200-5. Epub 2010 Feb 24. Semin Immunopathol. 2010. PMID: 20179931 Review.
-
Dominant role of antigen dose in CD4+Foxp3+ regulatory T cell induction and expansion.J Immunol. 2009 Oct 15;183(8):4895-903. doi: 10.4049/jimmunol.0901459. J Immunol. 2009. PMID: 19801514 Free PMC article.
Cited by
-
Soluble CD25 imposes a low-zone IL-2 signaling environment that favors competitive outgrowth of antigen-experienced CD25high regulatory and memory T cells.Cell Immunol. 2023 Feb;384:104664. doi: 10.1016/j.cellimm.2023.104664. Epub 2023 Jan 5. Cell Immunol. 2023. PMID: 36642016 Free PMC article.
-
Cell and biomaterial delivery strategies to induce immune tolerance.Adv Drug Deliv Rev. 2023 Dec;203:115141. doi: 10.1016/j.addr.2023.115141. Epub 2023 Nov 18. Adv Drug Deliv Rev. 2023. PMID: 37980950 Free PMC article. Review.
-
New insights on the role of human leukocyte antigen complex in primary biliary cholangitis.Front Immunol. 2022 Aug 31;13:975115. doi: 10.3389/fimmu.2022.975115. eCollection 2022. Front Immunol. 2022. PMID: 36119102 Free PMC article. Review.
-
Opposite Effects of mRNA-Based and Adenovirus-Vectored SARS-CoV-2 Vaccines on Regulatory T Cells: A Pilot Study.Biomedicines. 2023 Feb 10;11(2):511. doi: 10.3390/biomedicines11020511. Biomedicines. 2023. PMID: 36831046 Free PMC article.
-
Low-Zone IL-2 Signaling: Fusion Proteins Containing Linked CD25 and IL-2 Domains Sustain Tolerogenic Vaccination in vivo and Promote Dominance of FOXP3+ Tregs in vitro.Front Immunol. 2020 Sep 23;11:541619. doi: 10.3389/fimmu.2020.541619. eCollection 2020. Front Immunol. 2020. PMID: 33072087 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
