

Strongly Bent Double-Stranded DNA: Reconciling Theory and Experiment
- PMID: 32601596
- PMCID: PMC7323118
- DOI: 10.3389/fphy.2019.00195
Strongly Bent Double-Stranded DNA: Reconciling Theory and Experiment
Abstract
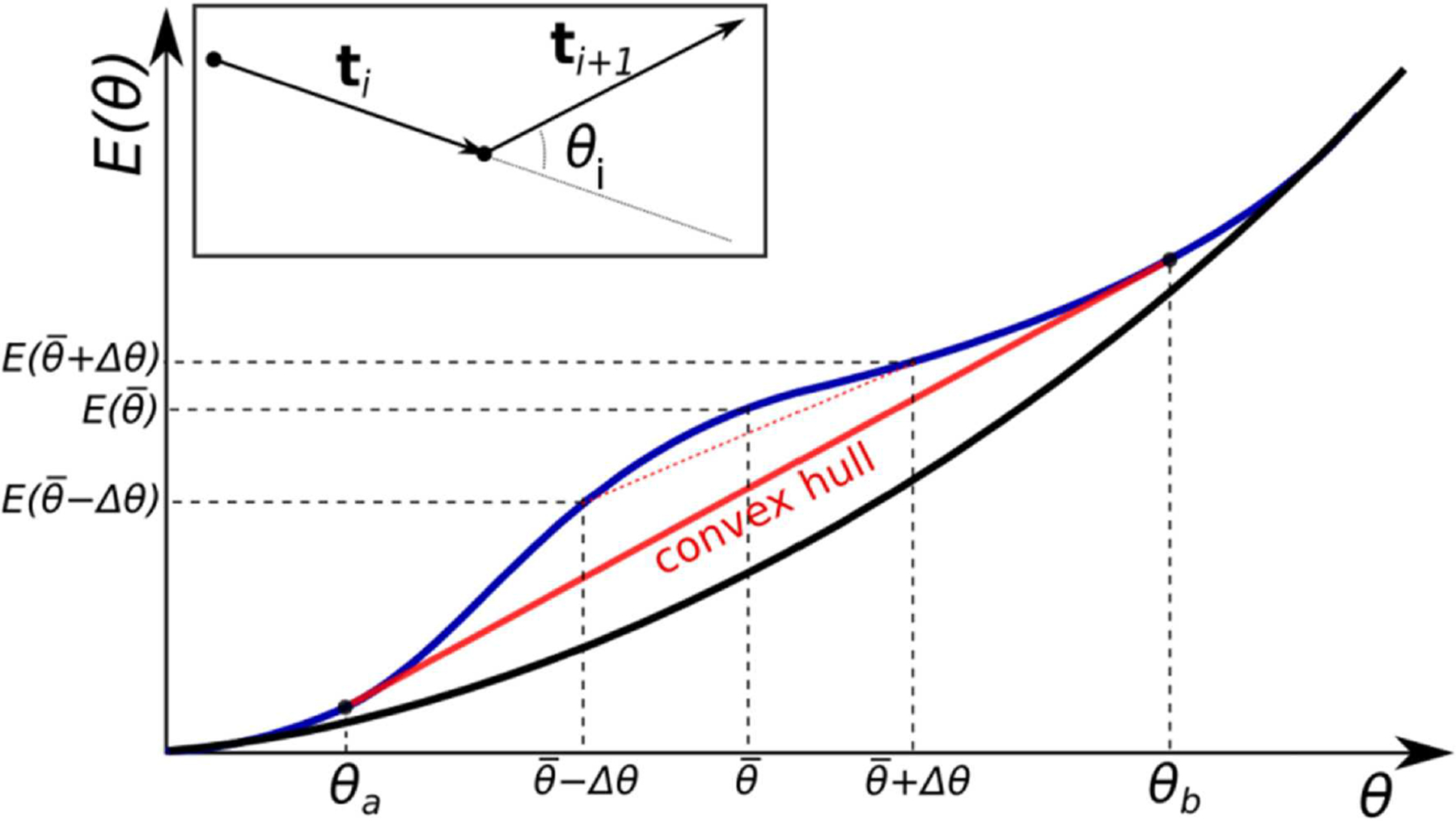
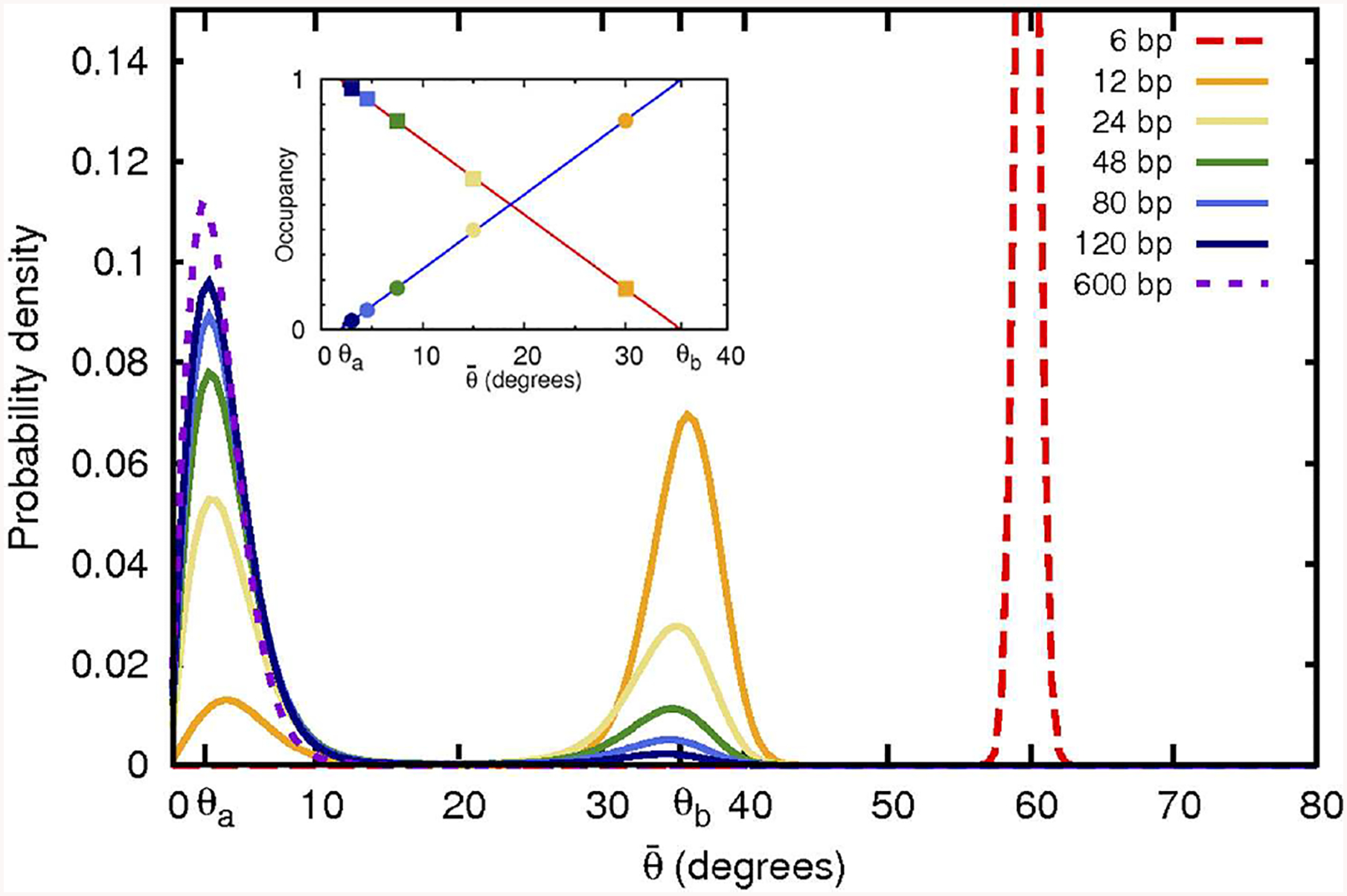
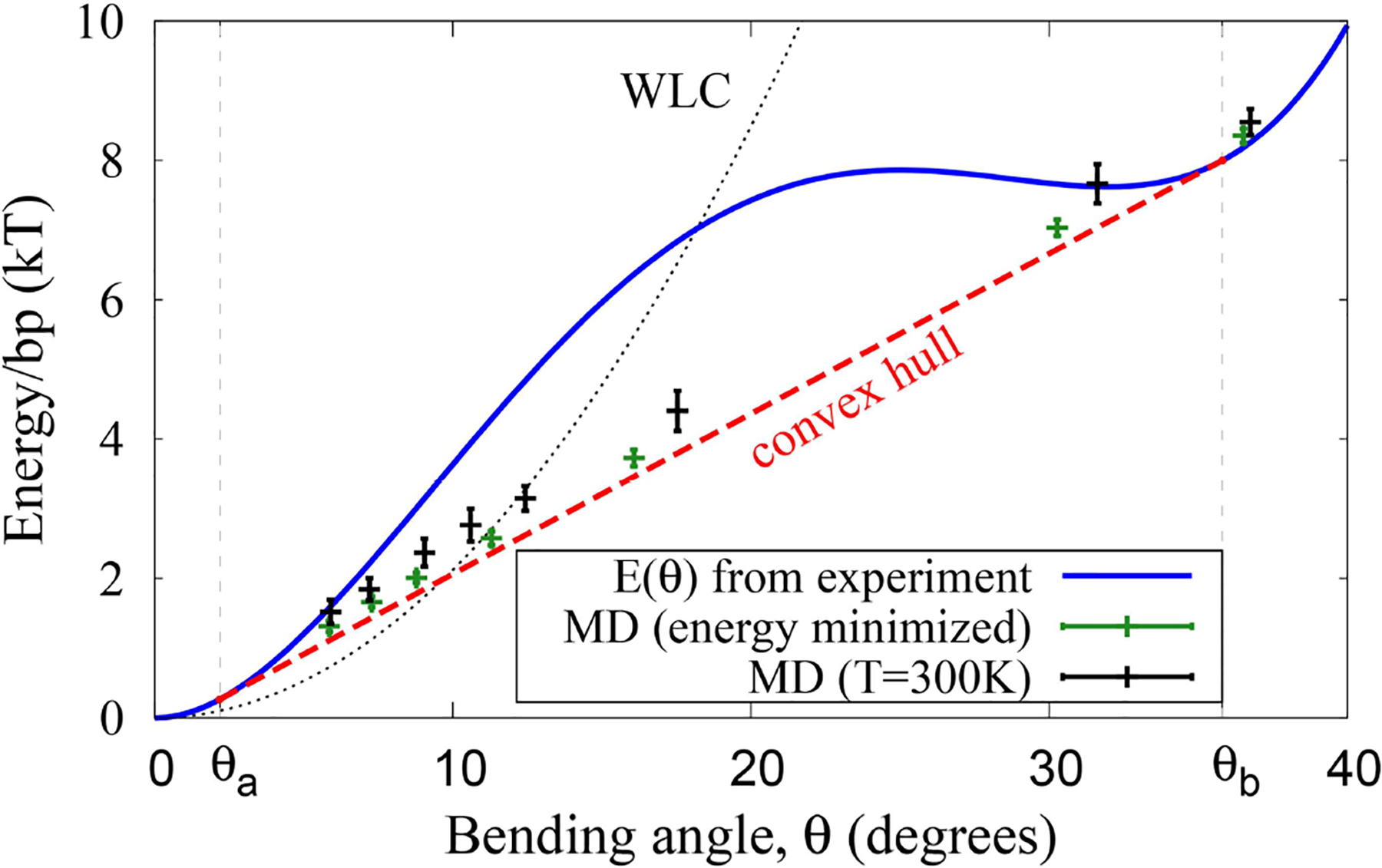
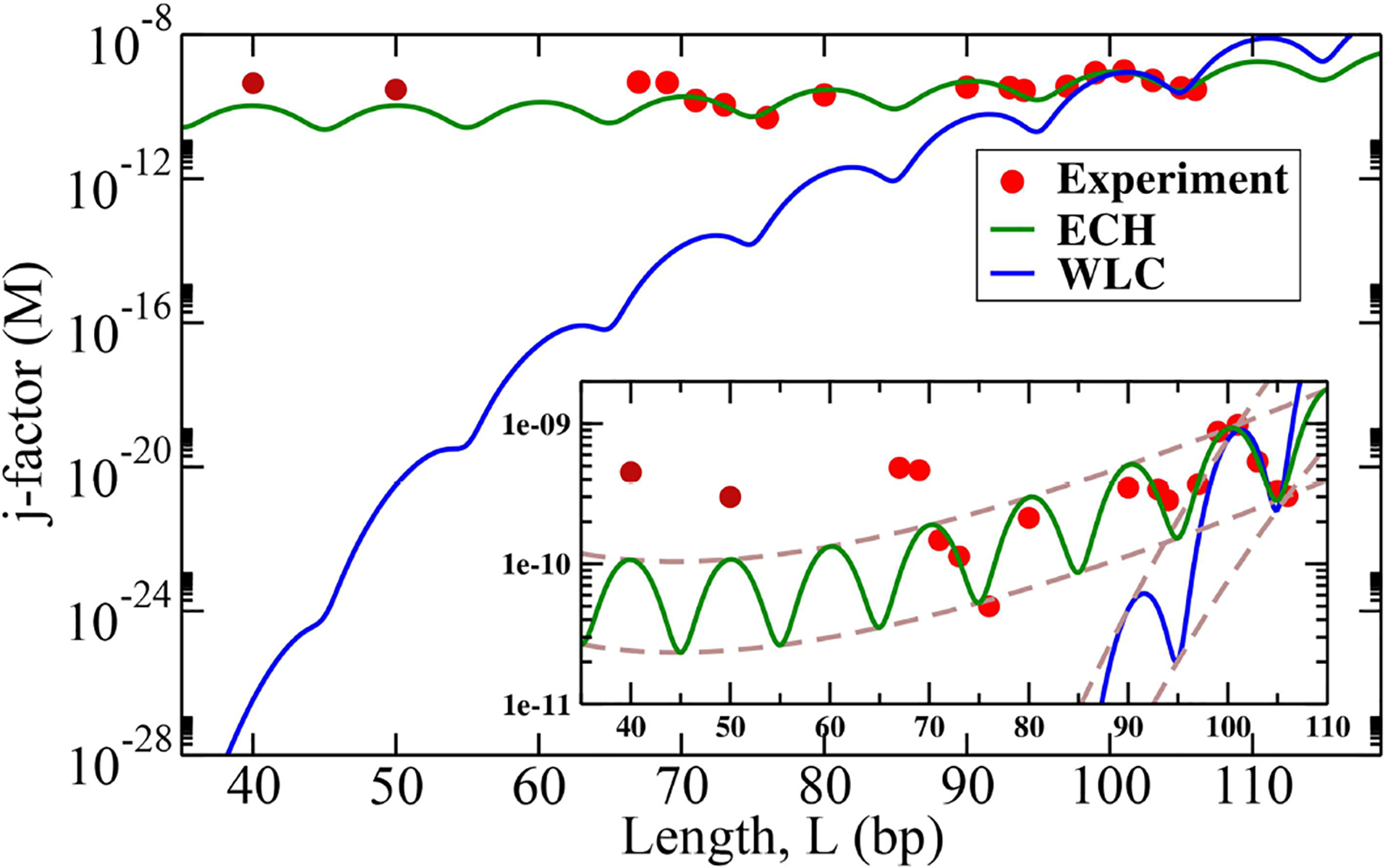
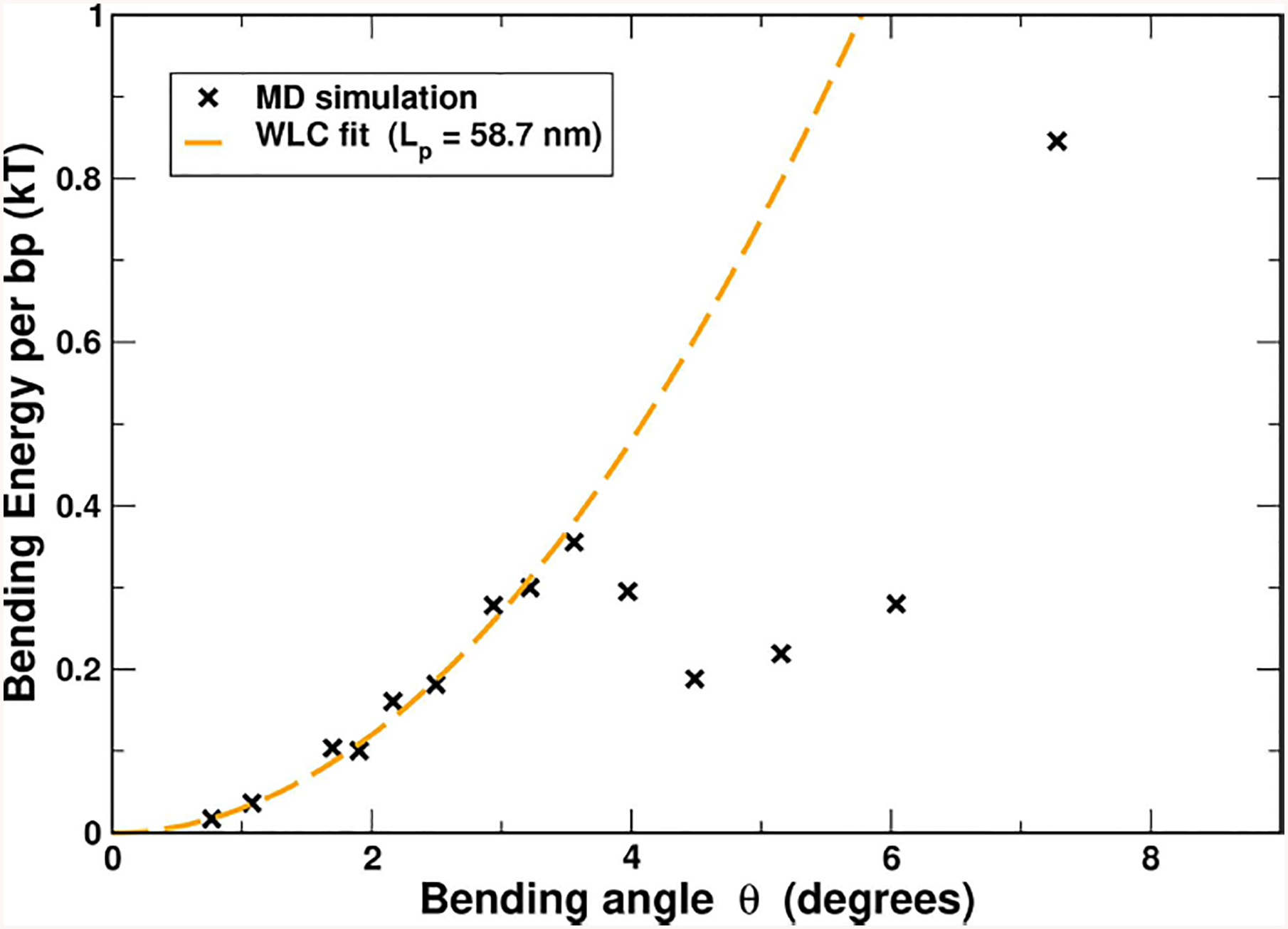
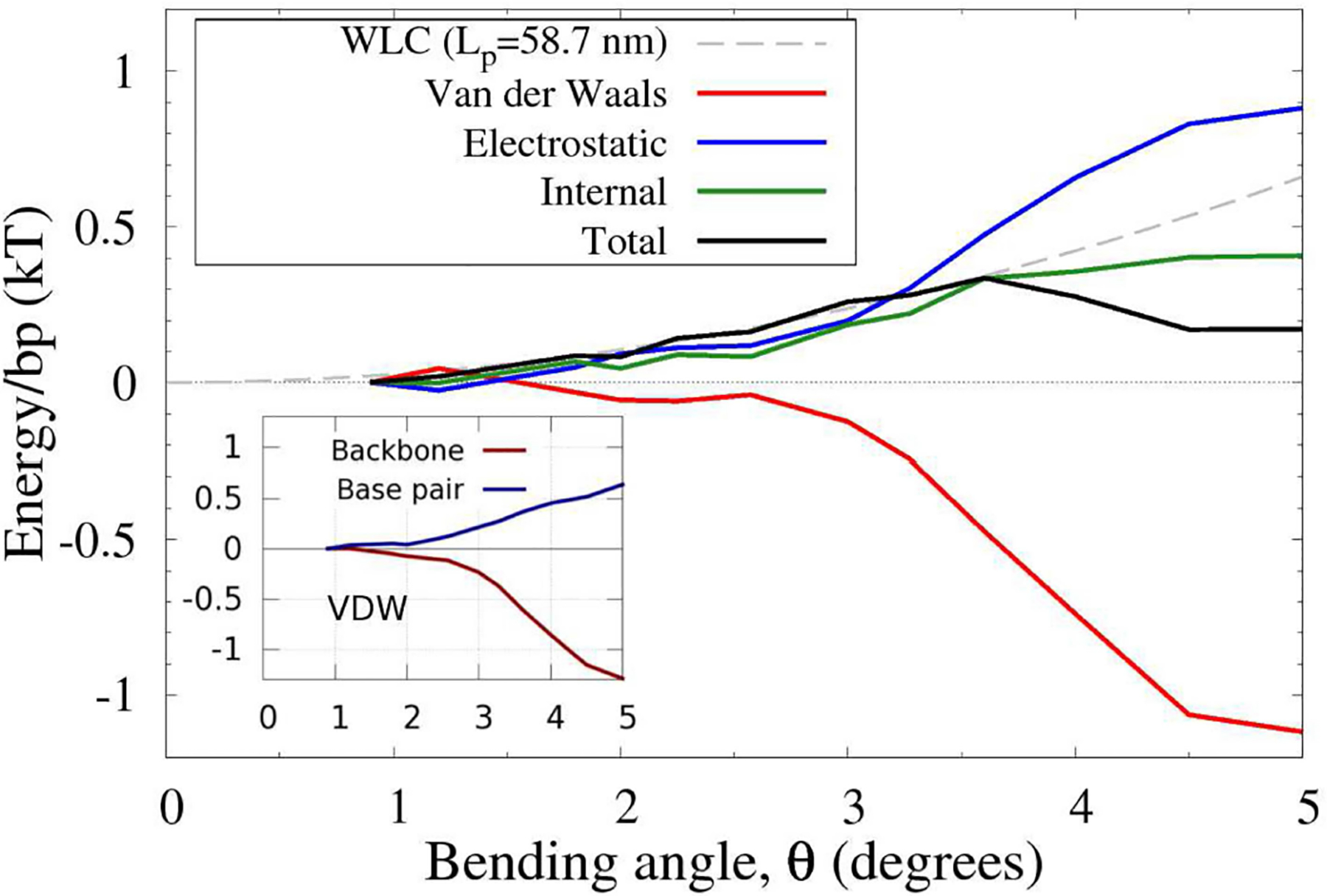
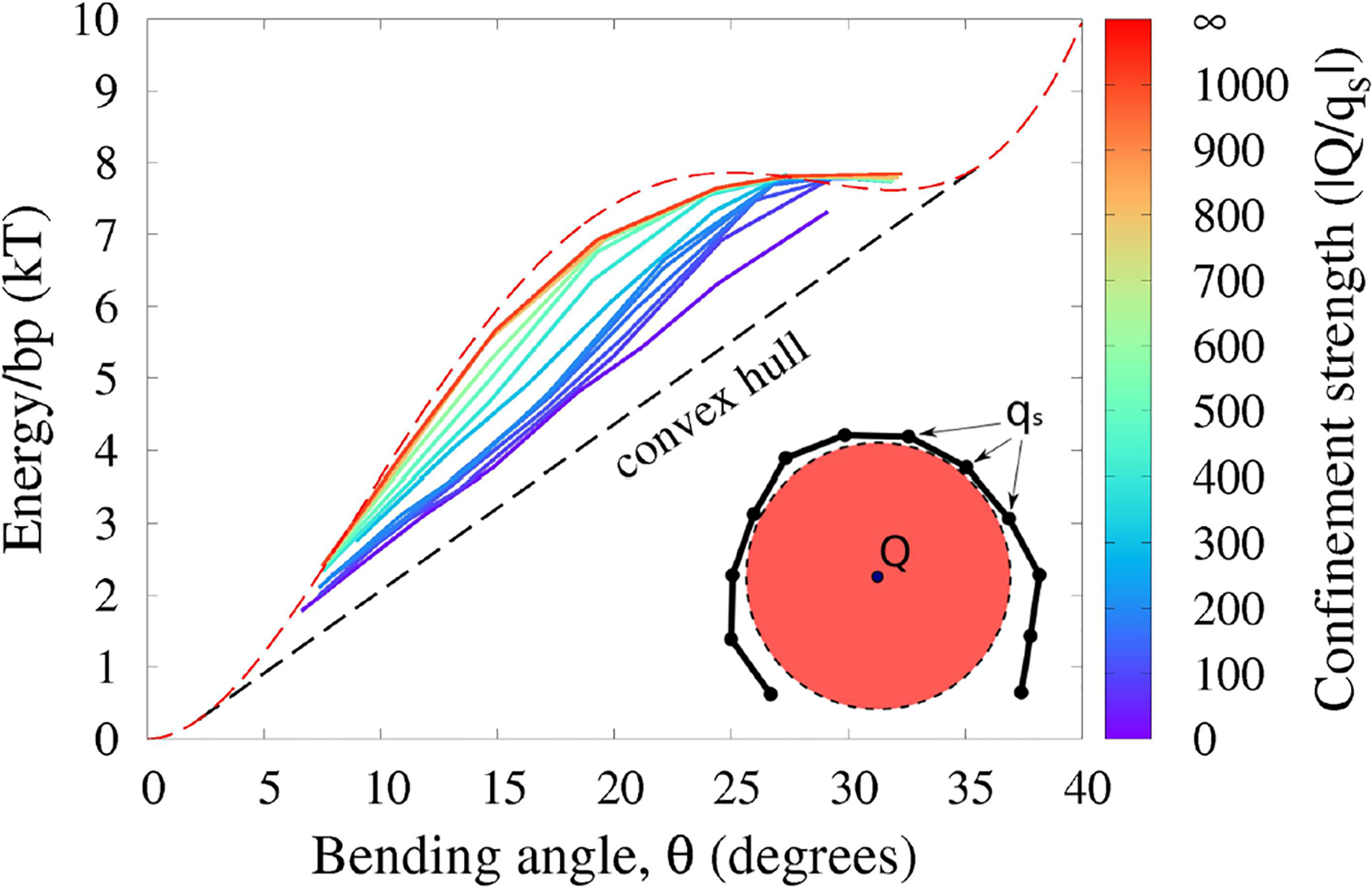
The strong bending of polymers is poorly understood. We propose a general quantitative framework of polymer bending that includes both the weak and strong bending regimes on the same footing, based on a single general physical principle. As the bending deformation increases beyond a certain (polymer-specific) point, the change in the convexity properties of the effective bending energy of the polymer makes the harmonic deformation energetically unfavorable: in this strong bending regime the energy of the polymer varies linearly with the average bending angle as the system follows the convex hull of the deformation energy function. For double-stranded DNA, the effective bending deformation energy becomes non-convex for bends greater than ~ 2° per base-pair, equivalent to the curvature of a closed circular loop of ~ 160 base pairs. A simple equation is derived for the polymer loop energy that covers both the weak and strong bending regimes. The theory shows quantitative agreement with recent DNA cyclization experiments on short DNA fragments, while maintaining the expected agreement with experiment in the weak bending regime. Counter-intuitively, cyclization probability (j-factor) of very short DNA loops is predicted to increase with decreasing loop length; the j-factor reaches its minimum for loops of ≃ 45 base pairs. Atomistic simulations reveal that the attractive component of the short-range Lennard-Jones interaction between the backbone atoms can explain the underlying non-convexity of the DNA effective bending energy, leading to the linear bending regime. Applicability of the theory to protein-DNA complexes, including the nucleosome, is discussed.
Keywords: DNA; convex hull; cyclization; deformation; j-factor; polymer bending.
Conflict of interest statement
Conflict of Interest: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
How a short double-stranded DNA bends.J Chem Phys. 2015 Apr 21;142(15):155101. doi: 10.1063/1.4916379. J Chem Phys. 2015. PMID: 25903911
-
Unwinding Induced Melting of Double-Stranded DNA Studied by Free Energy Simulations.J Phys Chem B. 2017 Dec 14;121(49):11019-11030. doi: 10.1021/acs.jpcb.7b07701. Epub 2017 Nov 19. J Phys Chem B. 2017. PMID: 29064703
-
Bending and circularization of site-specific and stereoisomeric carcinogen-DNA adducts.Biochemistry. 1998 Jan 13;37(2):769-78. doi: 10.1021/bi971785x. Biochemistry. 1998. PMID: 9425101
-
Packaged DNA. An elastic model.Cell Biophys. 1985 Mar;7(1):57-89. doi: 10.1007/BF02788639. Cell Biophys. 1985. PMID: 2408756 Review.
-
Two distinct modes of protein-induced bending in DNA.J Mol Biol. 1998 Sep 18;282(2):331-43. doi: 10.1006/jmbi.1998.1994. J Mol Biol. 1998. PMID: 9735291 Review.
Cited by
-
The greatest contribution to medical science is the transformation from studying symptoms to studying their causes-the unrelenting legacy of Robert Koch and Louis Pasteur-and a causality perspective to approach a definition of SLE.Front Immunol. 2024 Feb 1;15:1346619. doi: 10.3389/fimmu.2024.1346619. eCollection 2024. Front Immunol. 2024. PMID: 38361929 Free PMC article.
-
Structural Landscape of the Transition from an ssDNA Dumbbell Plus Its Complementary Hairpin to a dsDNA Microcircle Via a Kissing Loop Intermediate.Molecules. 2021 May 19;26(10):3017. doi: 10.3390/molecules26103017. Molecules. 2021. PMID: 34069399 Free PMC article.
-
Ion-mediated interactions between like-charged polyelectrolytes with bending flexibility.Sci Rep. 2020 Dec 9;10(1):21586. doi: 10.1038/s41598-020-78684-6. Sci Rep. 2020. PMID: 33299024 Free PMC article.
-
The Anti-DNA Antibodies: Their Specificities for Unique DNA Structures and Their Unresolved Clinical Impact-A System Criticism and a Hypothesis.Front Immunol. 2022 Jan 11;12:808008. doi: 10.3389/fimmu.2021.808008. eCollection 2021. Front Immunol. 2022. PMID: 35087528 Free PMC article. Review.
-
Experimental study of a nanoscale translocation ratchet.Proc Natl Acad Sci U S A. 2022 Jul 26;119(30):e2202527119. doi: 10.1073/pnas.2202527119. Epub 2022 Jul 18. Proc Natl Acad Sci U S A. 2022. PMID: 35858428 Free PMC article.
References
-
- Grosberg AY, Khokhlov AR, Jelinski LW. Giant molecules: here, there, and everywhere. Am J Phys. (1997) 65:1218–9.
-
- Van de Velde K, Kiekens P. Biopolymers: overview of several properties and consequences on their applications. Polym Test. (2002) 21:433–42. doi: 10.1016/S0142-9418(01)00107-6 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials