

Mechanisms of procollagen and HSP47 sorting during ER-to-Golgi trafficking
- PMID: 32562852
- PMCID: PMC8932071
- DOI: 10.1016/j.matbio.2020.06.002
Mechanisms of procollagen and HSP47 sorting during ER-to-Golgi trafficking
Abstract
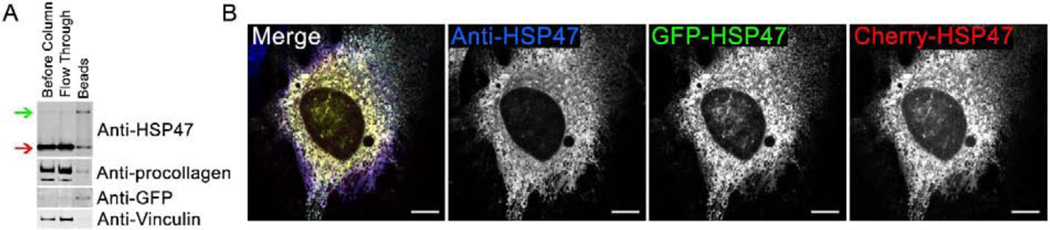
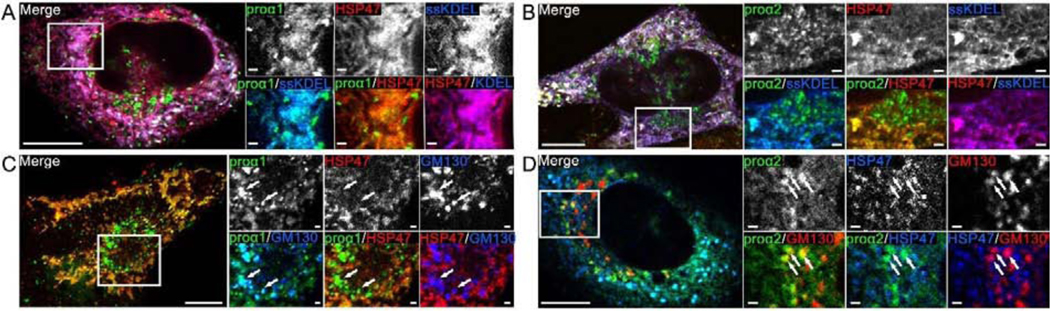
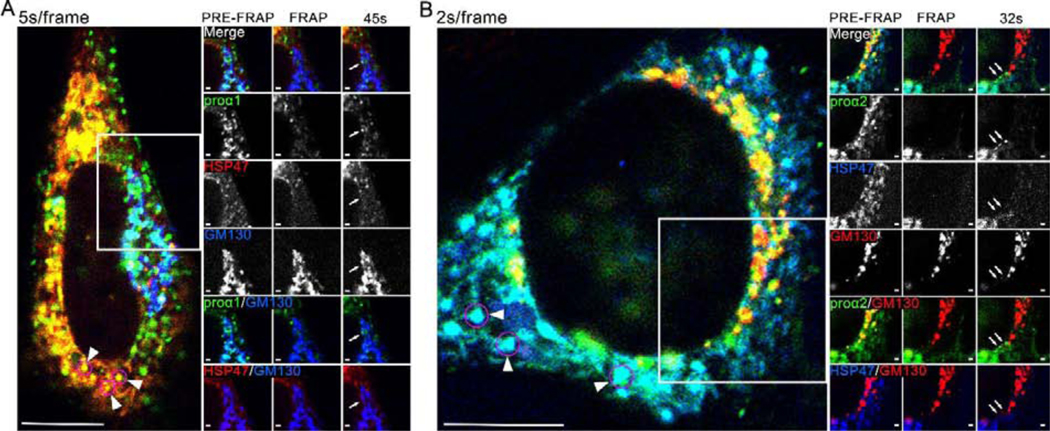
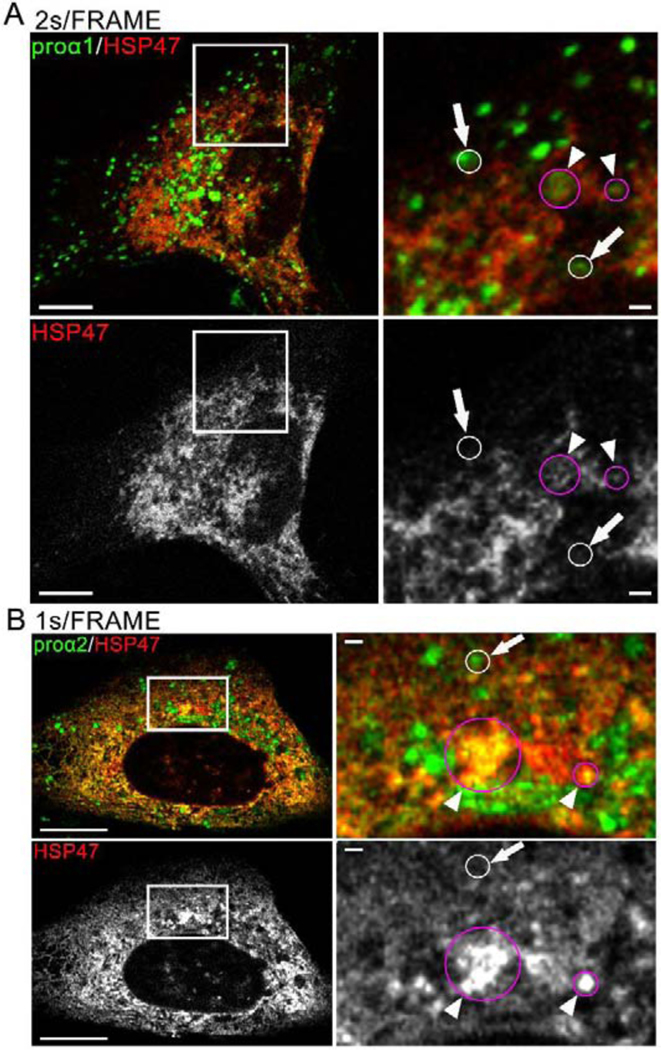
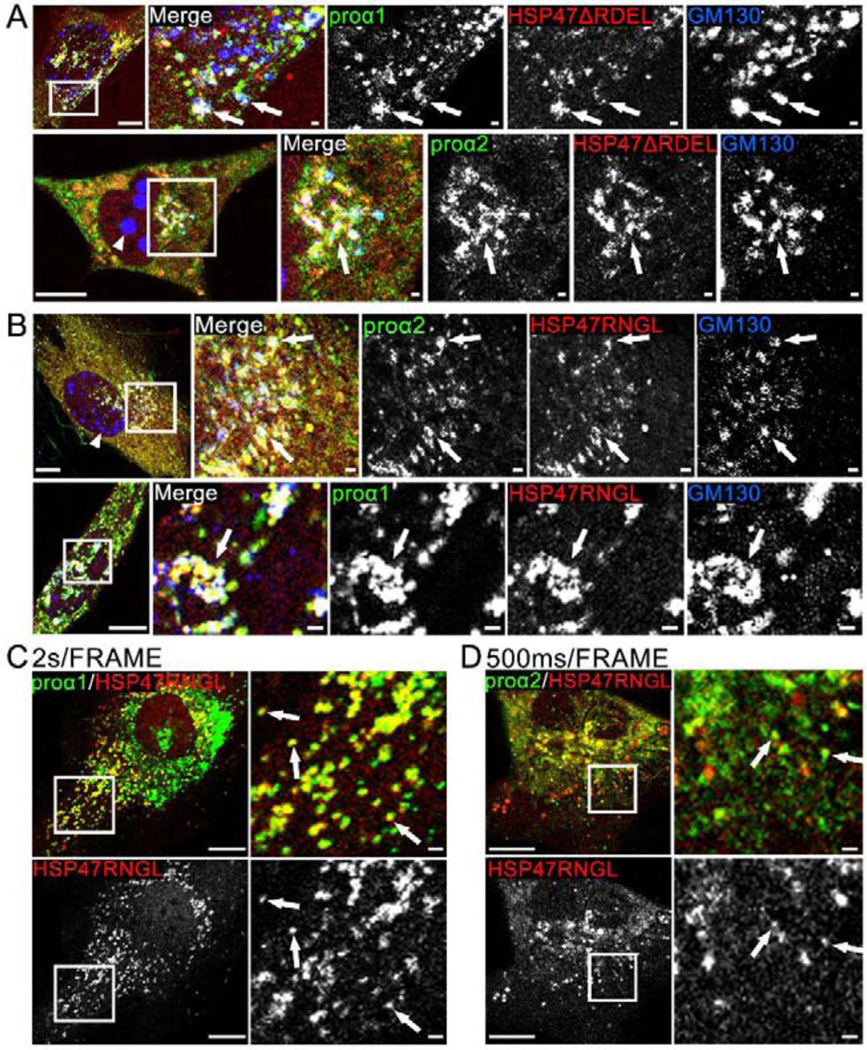
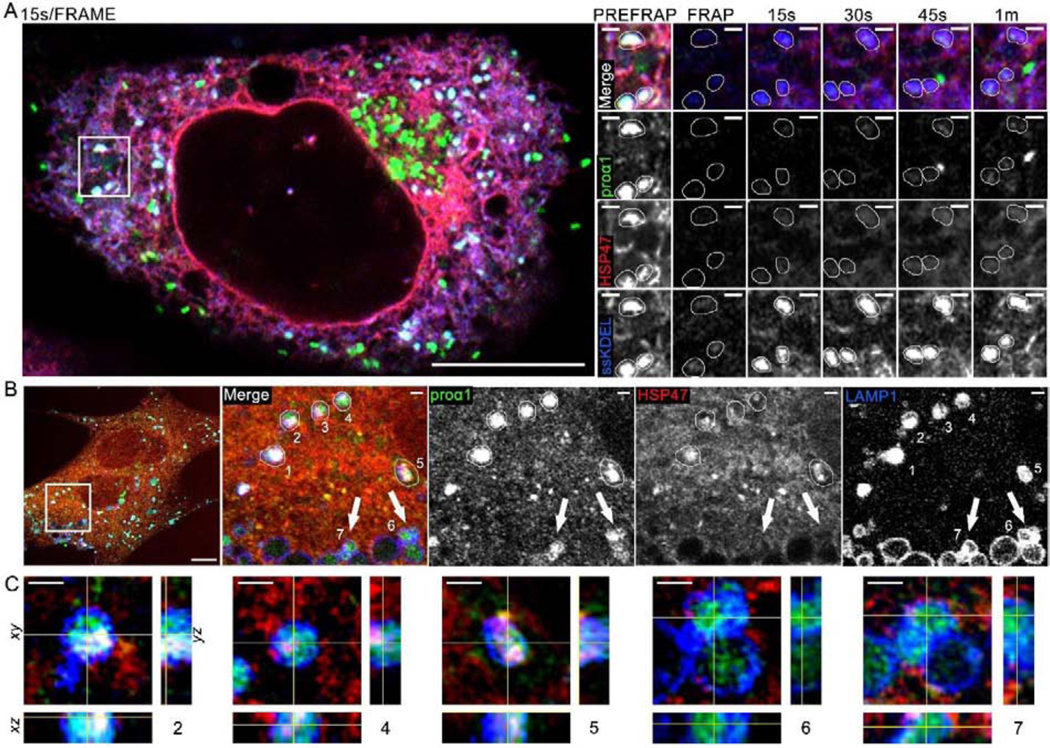
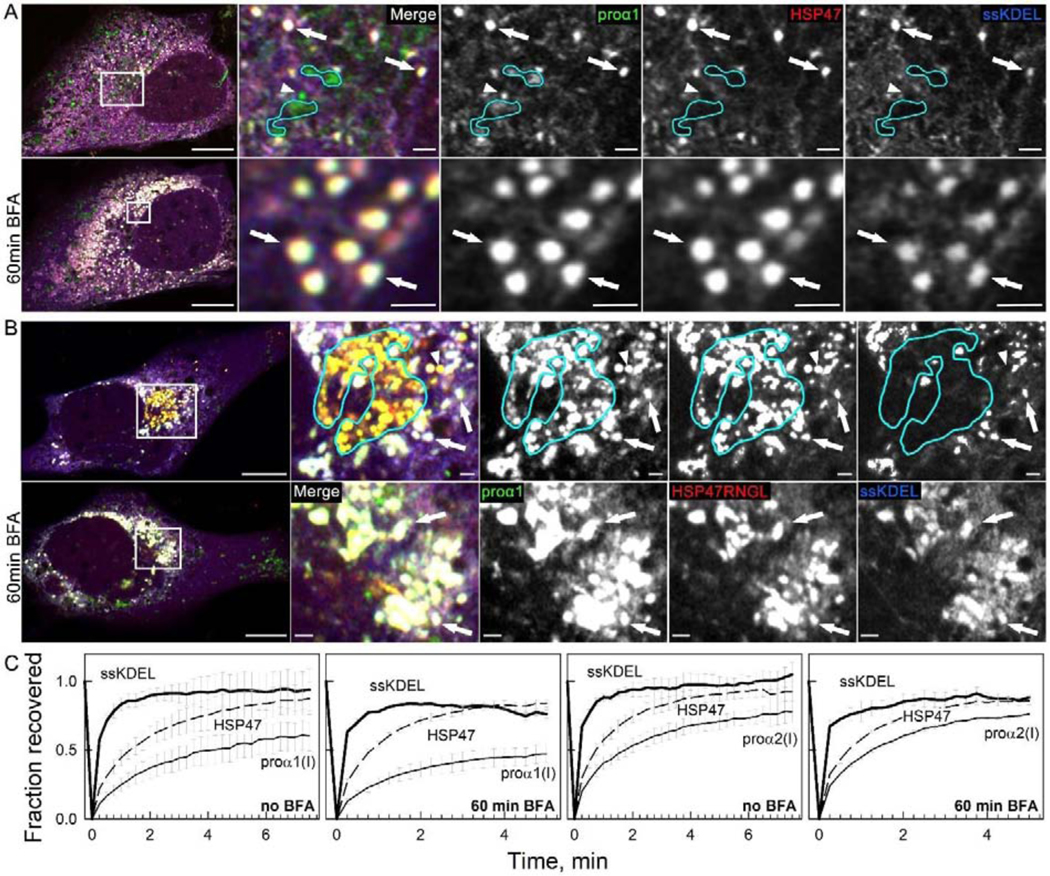
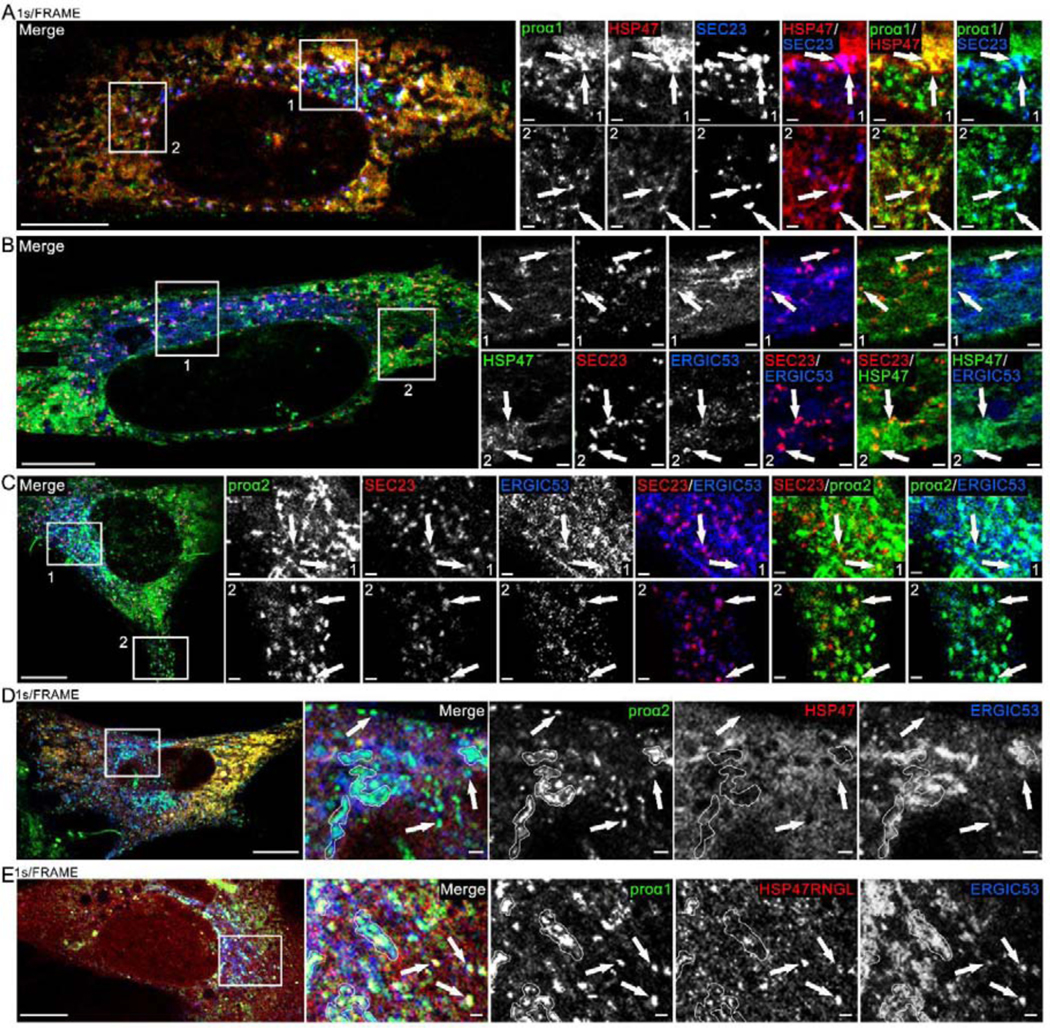
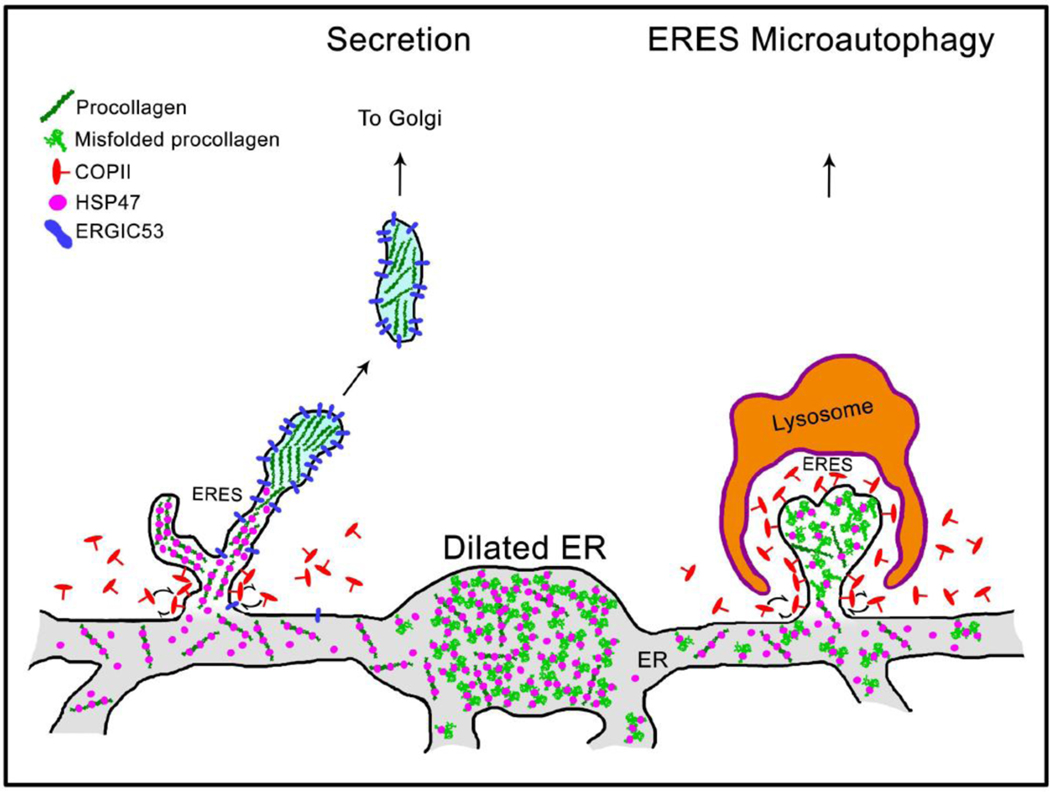
Efficient quality control and export of procollagen from the cell is crucial for extracellular matrix homeostasis, yet it is still incompletely understood. One of the debated questions is the role of a collagen-specific ER chaperone HSP47 in these processes. Most ER chaperones preferentially bind to unfolded polypeptide chains, enabling selective export of natively folded proteins from the ER after chaperone release. In contrast, HSP47 preferentially binds to the natively folded procollagen and is believed to be released only in the ER-Golgi intermediate compartment (ERGIC) or cis-Golgi. HSP47 colocalization with procollagen in punctate structures observed by immunofluorescence imaging of fixed cells has thus been interpreted as evidence for HSP47 export from the ER together with procollagen in transport vesicles destined for ERGIC or Golgi. To understand the mechanism of this co-trafficking and its physiological significance, we imaged the dynamics of fluorescently tagged type I procollagen and HSP47 punctate structures in live MC3T3 murine osteoblasts with up to 120 nm spatial and 500 ms time resolution. Contrary to the prevailing model, we discovered that most bona fide carriers delivering procollagen from ER exit sites (ERESs) to Golgi contained no HSP47, unless the RDEL signal for ER retention in HSP47 was deleted or mutated. These transport intermediates exhibited characteristic rapid, directional motion along microtubules, while puncta with colocalized HSP47 and procollagen similar to the ones described before had only limited, stochastic motion. Live cell imaging and fluorescence recovery after photobleaching revealed that the latter puncta (including the ones induced by ARF1 inhibition) were dilated regions of ER lumen, ERESs, or autophagic structures surrounded by lysosomal membranes. Procollagen was colocalized with HSP47 and ERGIC53 at ERESs. It was colocalized with ERGIC53 but not HSP47 in Golgi-bound transport intermediates. Our results suggest that procollagen and HSP47 sorting occurs at ERES before procollagen is exported from the ER in Golgi-bound transport intermediates, providing new insights into mechanisms of procollagen trafficking.
Keywords: Collagen; ER exit sites; HSP47; live cell imaging; trafficking.
Published by Elsevier B.V.
Conflict of interest statement
Declaration of Competing Interests The authors have no competing interests.
Figures
Similar articles
-
Noncanonical ER-Golgi trafficking and autophagy of endogenous procollagen in osteoblasts.Cell Mol Life Sci. 2021 Dec;78(24):8283-8300. doi: 10.1007/s00018-021-04017-z. Epub 2021 Nov 15. Cell Mol Life Sci. 2021. PMID: 34779895 Free PMC article.
-
Hsp47 and cyclophilin B traverse the endoplasmic reticulum with procollagen into pre-Golgi intermediate vesicles. A role for Hsp47 and cyclophilin B in the export of procollagen from the endoplasmic reticulum.J Biol Chem. 1995 Aug 4;270(31):18323-8. doi: 10.1074/jbc.270.31.18323. J Biol Chem. 1995. PMID: 7629154
-
Intracellular interaction of collagen-specific stress protein HSP47 with newly synthesized procollagen.J Cell Biol. 1996 Apr;133(2):469-83. doi: 10.1083/jcb.133.2.469. J Cell Biol. 1996. PMID: 8609177 Free PMC article.
-
Expression and function of heat shock protein 47: a collagen-specific molecular chaperone in the endoplasmic reticulum.Matrix Biol. 1998 Feb;16(7):379-86. doi: 10.1016/s0945-053x(98)90011-7. Matrix Biol. 1998. PMID: 9524358 Review.
-
Roles of the endoplasmic reticulum-resident, collagen-specific molecular chaperone Hsp47 in vertebrate cells and human disease.J Biol Chem. 2019 Feb 8;294(6):2133-2141. doi: 10.1074/jbc.TM118.002812. Epub 2018 Dec 12. J Biol Chem. 2019. PMID: 30541925 Free PMC article. Review.
Cited by
-
TANGO1 Dances to Export of Procollagen from the Endoplasmic Reticulum.Fibrosis (Hong Kong). 2023 Dec;1(2):10008. doi: 10.35534/fibrosis.2023.10008. Epub 2023 Dec 21. Fibrosis (Hong Kong). 2023. PMID: 38650832 Free PMC article.
-
Genetically encoded fluorescent tools: Shining a little light on ER-to-Golgi transport.Free Radic Biol Med. 2022 Apr;183:14-24. doi: 10.1016/j.freeradbiomed.2022.03.004. Epub 2022 Mar 8. Free Radic Biol Med. 2022. PMID: 35272000 Free PMC article. Review.
-
Autophagy guards tendon homeostasis.Cell Death Dis. 2022 Apr 23;13(4):402. doi: 10.1038/s41419-022-04824-7. Cell Death Dis. 2022. PMID: 35461310 Free PMC article.
-
ER-phagy: mechanisms, regulation, and diseases connected to the lysosomal clearance of the endoplasmic reticulum.Physiol Rev. 2022 Jul 1;102(3):1393-1448. doi: 10.1152/physrev.00038.2021. Epub 2022 Feb 21. Physiol Rev. 2022. PMID: 35188422 Free PMC article. Review.
-
Targeting cellular stress in vitro improves osteoblast homeostasis, matrix collagen content and mineralization in two murine models of osteogenesis imperfecta.Matrix Biol. 2021 Apr;98:1-20. doi: 10.1016/j.matbio.2021.03.001. Epub 2021 Mar 31. Matrix Biol. 2021. PMID: 33798677 Free PMC article.
References
-
- McCaughey J, Stephens DJ, ER-to-Golgi Transport: A Sizeable Problem, Trends Cell Biol 29(12) (2019) 940–953. - PubMed
-
- McGourty CA, Akopian D, Walsh C, Gorur A, Werner A, Schekman R, Bautista D, Rape M, Regulation of the CUL3 Ubiquitin Ligase by a Calcium-Dependent Co-adaptor, Cell 167(2) (2016) 525–538 e14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
