

Non-Mouse-Adapted H1N1pdm09 Virus as a Model for Influenza Research
- PMID: 32485821
- PMCID: PMC7354452
- DOI: 10.3390/v12060590
Non-Mouse-Adapted H1N1pdm09 Virus as a Model for Influenza Research
Abstract
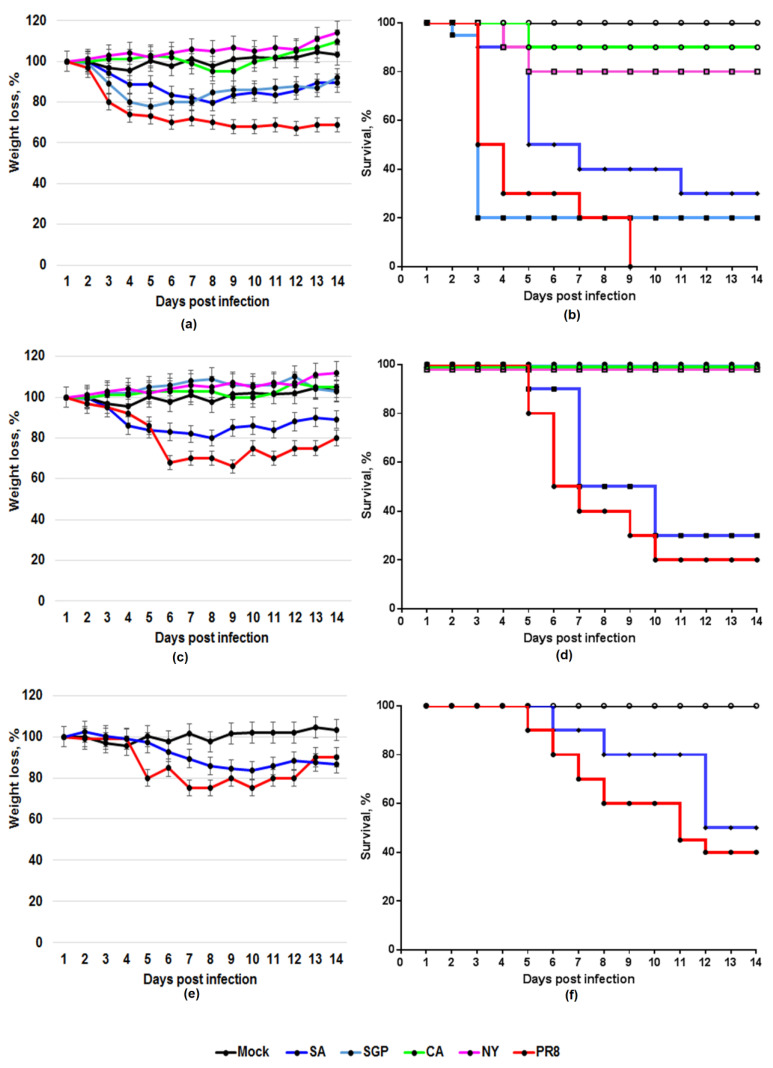
The number of lung-adapted influenza viruses is limited. Most of them are not antigenically related to current circulating viruses. Viruses similar to recent strains are required for screening modern antiviral compounds and studying new vaccine candidates against novel influenza viruses. The process by which an influenza virus adapts to a new host is rather difficult. The aim of this study was to select a non-adapted current virus whose major biological properties correspond to those of classical lab-adapted viruses. Mice were inoculated intranasally with non-lung-adapted influenza viruses of subtype H1N1pdm09. They were monitored closely for body weight loss, mortality outcomes and gross pathology for 14 days following inoculation, as well as viral replication in lung tissue. Lung-adapted PR8 virus was used as a control. The tested viruses multiplied equally well in the lower respiratory tract of mice without prior adaptation but dramatically differed in lethality; the differences in their toxicity and pathogenicity in mice were established. A/South Africa/3626/2013 (H1N1)pdm09 virus was found to be an appropriate candidate to replace PR8 as a model virus for influenza research. No prior adaptation to the animal model is needed to reach the pathogenicity level of the classical mouse-adapted PR8 virus.
Keywords: animal model; influenza infection; influenza virus; viral pathogenesis; viral toxicity.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
Comparative In Vitro and In Vivo Analysis of H1N1 and H1N2 Variant Influenza Viruses Isolated from Humans between 2011 and 2016.J Virol. 2018 Oct 29;92(22):e01444-18. doi: 10.1128/JVI.01444-18. Print 2018 Nov 15. J Virol. 2018. PMID: 30158292 Free PMC article.
-
An early 'classical' swine H1N1 influenza virus shows similar pathogenicity to the 1918 pandemic virus in ferrets and mice.Virology. 2009 Oct 25;393(2):338-45. doi: 10.1016/j.virol.2009.08.021. Epub 2009 Sep 5. Virology. 2009. PMID: 19733889 Free PMC article.
-
Hemagglutinin Stability Regulates H1N1 Influenza Virus Replication and Pathogenicity in Mice by Modulating Type I Interferon Responses in Dendritic Cells.J Virol. 2020 Jan 17;94(3):e01423-19. doi: 10.1128/JVI.01423-19. Print 2020 Jan 17. J Virol. 2020. PMID: 31694942 Free PMC article.
-
The contribution of animal models to the understanding of the host range and virulence of influenza A viruses.Microbes Infect. 2011 May;13(5):502-15. doi: 10.1016/j.micinf.2011.01.014. Epub 2011 Jan 27. Microbes Infect. 2011. PMID: 21276869 Free PMC article. Review.
-
Nonrespiratory sites of influenza-associated disease: mechanisms and experimental systems for continued study.FEBS J. 2022 Jul;289(14):4038-4060. doi: 10.1111/febs.16363. Epub 2022 Feb 7. FEBS J. 2022. PMID: 35060315 Free PMC article. Review.
Cited by
-
4'-Fluorouridine mitigates lethal infection with pandemic human and highly pathogenic avian influenza viruses.PLoS Pathog. 2023 Apr 17;19(4):e1011342. doi: 10.1371/journal.ppat.1011342. eCollection 2023 Apr. PLoS Pathog. 2023. PMID: 37068076 Free PMC article.
-
Universal Live-Attenuated Influenza Vaccine Candidates Expressing Multiple M2e Epitopes Protect Ferrets against a High-Dose Heterologous Virus Challenge.Viruses. 2021 Jun 30;13(7):1280. doi: 10.3390/v13071280. Viruses. 2021. PMID: 34209093 Free PMC article.
-
Anti-Influenza Effect of Nanosilver in a Mouse Model.Vaccines (Basel). 2020 Nov 13;8(4):679. doi: 10.3390/vaccines8040679. Vaccines (Basel). 2020. PMID: 33202939 Free PMC article.
-
Understanding the Variability of Certain Biological Properties of H1N1pdm09 Influenza Viruses.Vaccines (Basel). 2022 Mar 3;10(3):395. doi: 10.3390/vaccines10030395. Vaccines (Basel). 2022. PMID: 35335027 Free PMC article.
-
The Effect of Mice Adaptation Process on the Pathogenicity of Influenza A/South Africa/3626/2013 (H1N1)pdm09 Model Strain.Int J Mol Sci. 2023 Dec 12;24(24):17386. doi: 10.3390/ijms242417386. Int J Mol Sci. 2023. PMID: 38139214 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
