

Review
doi: 10.1038/s41577-020-0285-6.
Epub 2020 Mar 4.
Defining trained immunity and its role in health and disease
Affiliations
- PMID: 32132681
- PMCID: PMC7186935
- DOI: 10.1038/s41577-020-0285-6
Item in Clipboard
Review
Defining trained immunity and its role in health and disease
Nat Rev Immunol.
2020 Jun.
Abstract
Immune memory is a defining feature of the acquired immune system, but activation of the innate immune system can also result in enhanced responsiveness to subsequent triggers. This process has been termed 'trained immunity', a de facto innate immune memory. Research in the past decade has pointed to the broad benefits of trained immunity for host defence but has also suggested potentially detrimental outcomes in immune-mediated and chronic inflammatory diseases. Here we define 'trained immunity' as a biological process and discuss the innate stimuli and the epigenetic and metabolic reprogramming events that shape the induction of trained immunity.
Conflict of interest statement
The authors declare no competing interests.
Figures
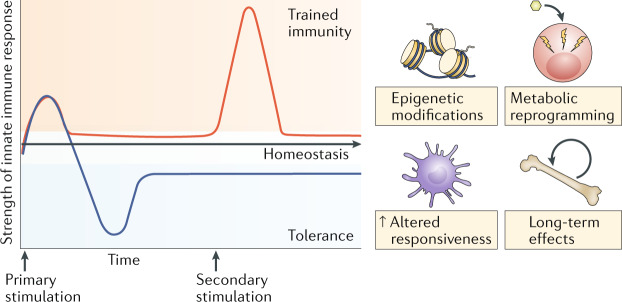
Infections or sterile tissue triggers induce inflammation and the activation of immune effector mechanisms. Concomitant to a proinflammatory response, anti-inflammatory mechanisms are provoked to prevent overshooting inflammation and tissue damage and to limit the inflammatory response in time. Trained immunity involves epigenetic and metabolic reprogramming of the innate immune cells, allowing qualitatively and quantitatively adjusted responses of innate immune cells to subsequent time-delayed heterologous stimulation. Misguided trained immunity responses can contribute to disease progression, resulting in either a chronic hyperinflammatory state or a persistent state of immunological tolerance, a mechanism that dampens the inflammatory response of the host to maintain homeostasis and prevent tissue damage and organ failure, with the subsequent risk of secondary infections and other diseases related to decreased activity of the immune system.
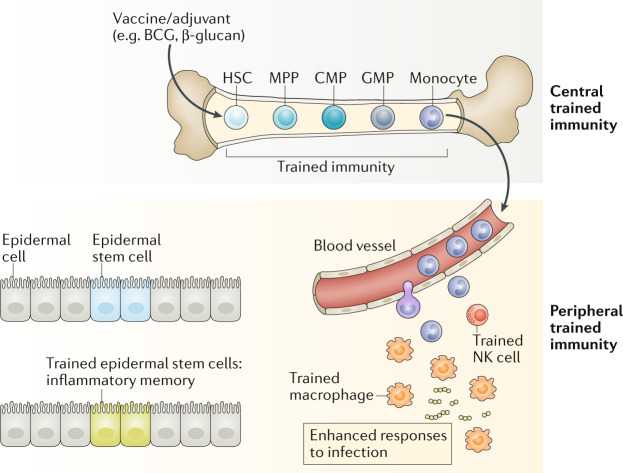
Although trained immunity was first established in cells of the mononuclear phagocyte lineage (that is, monocytes and macrophages), monocytes have a relatively short lifespan and are unlikely to transmit their memory phenotype to their progeny and provide sustainable protection. Thus, current vaccine strategies that directly target monocytes or macrophages may have limited capacity for generating sustained innate immune memory. By contrast, haematopoietic stem cells (HSCs) are long-lived cells with self-renewal properties that reside in the bone marrow. The bone marrow is the site of haematopoiesis where HSCs continually undergo asymmetric division giving rise to the full repertoire of myeloid and lymphoid cell types. HSCs can directly respond to acute and chronic infections. Although the exact mechanisms of precursor proliferation or differentiation are not well understood, persistent activation of HSCs can result in their exhaustion, leading to devastating effects on the systemic immune compartment. Monocytes derived from trained HSCs migrate to peripheral organs, where they give rise to monocyte-derived macrophages with enhanced effector functions against different types of pathogens. Natural killer (NK) cells possess adaptive immune characteristics following infection. On reinfection, these memory NK cells undergo a secondary expansion and can more rapidly degranulate and release cytokines, resulting in a more protective immune response. Epithelial stem cells show memory functions during human allergic inflammatory disease, displaying changes in the chromatin accessibility when the stimulus is withdrawn. BCG, bacillus Calmette–Guérin; CMP, common myeloid progenitor: GMP, granulocyte–macrophage progenitor; MPP, multipotent progenitor.
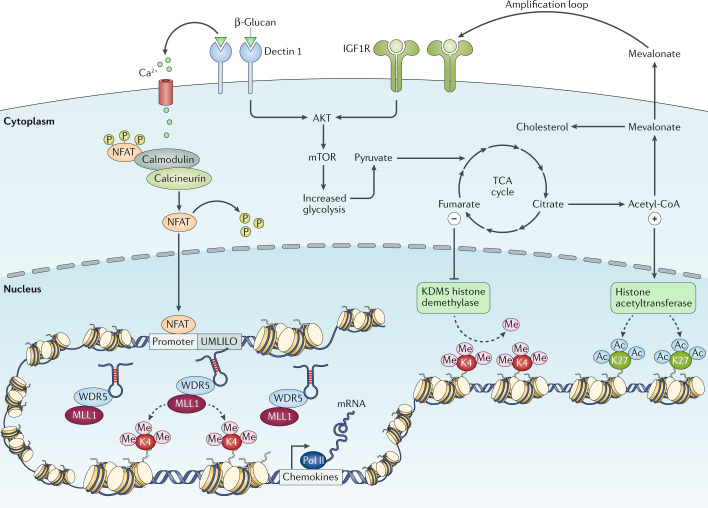
The correct initiation of the mechanisms necessary for the induction of trained immunity relies on the active interplay between epigenetic and metabolic reprogramming of the innate immune cells on stimulation. During primary challenge, the recognition of specific ligands by pattern recognition receptors triggers a series of intracellular cascades that lead to the upregulation of different metabolic pathways, such as glycolysis, tricarboxylic acid (TCA) cycle and fatty acid metabolism. Certain metabolites derived from these processes, such as fumarate and acetyl coenzyme A (acetyl-CoA), can activate or inhibit a series of enzymes involved in remodelling the epigenetic landscape of cells, such as the histone demethylase lysine-specific demethylase 5 (KDM5) or histone acetyltransferases, leading to specific changes in histone methylation and acetylation of genes involved in the innate immune responses. β-Glucan-mediated activation of dectin 1 signalling also triggers calcium influx, which leads to the dephosphorylation of nuclear factor of activated T cells (NFAT), allowing its translocation into the nucleus, where it may bind to DNA and activate gene transcription. This facilitates the accessibility of the DNA to the transcriptional machinery and gene regulatory elements and specific long non-coding RNAs, promoting and facilitating an enhanced gene transcription on secondary stimulation of the cells. IGF1R, insulin-like growth factor 1 receptor; MLL1, mixed-lineage leukaemia protein 1 (also known as histone-lysine N-methyltransferase 2A); mTOR, mechanistic target of rapamycin; Pol, polymerase; UMLILO, upstream master long non-coding RNA of the inflammatory chemokine locus; WDR5, WD repeat-containing protein 5.
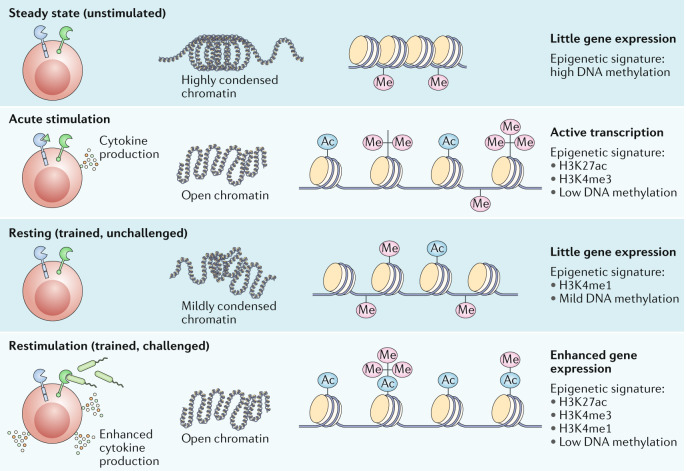
Stimulation of innate immune cells is accompanied by the deposition of chromatin marks and changes in the DNA methylation status, leading to unfolding of chromatin and facilitating transcription and expression of proinflammatory factors. All of these changes are only partially removed after cessation of the stimulus. This allows quicker and enhanced recruitment of transcription factors and gene expression after secondary challenge with another stimulus. The figure illustrates the chromatin states and epigenetic signatures associated with unstimulated cells, with cells following acute stimulation, with resting ‘trained’ cells and with trained cells following restimulation. H3K27ac, histone 3 lysine 27 acetylation; H3K4me, histone 3 lysine 4 methylation; H3K4me3, histone 3 lysine 4 trimethylation.
Similar articles
-
Trained immunity and host-pathogen interactions.Cell Microbiol. 2020 Dec;22(12):e13261. doi: 10.1111/cmi.13261. Epub 2020 Sep 17. Cell Microbiol. 2020. PMID: 32902895 Free PMC article. Review.
-
The Intersection of Epigenetics and Metabolism in Trained Immunity.Immunity. 2021 Jan 12;54(1):32-43. doi: 10.1016/j.immuni.2020.10.011. Epub 2020 Nov 20. Immunity. 2021. PMID: 33220235 Review.
-
Training innate immunity: the changing concept of immunological memory in innate host defence.Eur J Clin Invest. 2013 Aug;43(8):881-4. doi: 10.1111/eci.12132. Eur J Clin Invest. 2013. PMID: 23869409 Review.
-
Therapeutic targeting of trained immunity.Nat Rev Drug Discov. 2019 Jul;18(7):553-566. doi: 10.1038/s41573-019-0025-4. Nat Rev Drug Discov. 2019. PMID: 30967658 Free PMC article. Review.
-
Molecular and Cellular Mechanisms Modulating Trained Immunity by Various Cell Types in Response to Pathogen Encounter.Front Immunol. 2021 Oct 4;12:745332. doi: 10.3389/fimmu.2021.745332. eCollection 2021. Front Immunol. 2021. PMID: 34671359 Free PMC article. Review.
Cited by
-
Antiviral potential of plant polysaccharide nanoparticles actuating non-specific immunity.Int J Biol Macromol. 2021 Jul 1;182:743-749. doi: 10.1016/j.ijbiomac.2021.03.135. Epub 2021 Apr 5. Int J Biol Macromol. 2021. PMID: 33831450 Free PMC article.
-
Current Progress in the Science of Novel Adjuvant Nano-Vaccine-Induced Protective Immune Responses.Pathogens. 2024 May 23;13(6):441. doi: 10.3390/pathogens13060441. Pathogens. 2024. PMID: 38921739 Free PMC article. Review.
-
Could the Induction of Trained Immunity by β-Glucan Serve as a Defense Against COVID-19?Front Immunol. 2020 Jul 14;11:1782. doi: 10.3389/fimmu.2020.01782. eCollection 2020. Front Immunol. 2020. PMID: 32760409 Free PMC article.
-
Safety and COVID-19 Symptoms in Individuals Recently Vaccinated with BCG: a Retrospective Cohort Study.Cell Rep Med. 2020 Aug 25;1(5):100073. doi: 10.1016/j.xcrm.2020.100073. Epub 2020 Aug 5. Cell Rep Med. 2020. PMID: 32838341 Free PMC article.
-
Innate immune imprints in SARS-CoV-2 Omicron variant infection convalescents.Signal Transduct Target Ther. 2022 Nov 16;7(1):377. doi: 10.1038/s41392-022-01237-y. Signal Transduct Target Ther. 2022. PMID: 36379915 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
