

Priming of Antiviral CD8 T Cells without Effector Function by a Persistently Replicating Hepatitis C-Like Virus
- PMID: 32102885
- PMCID: PMC7199419
- DOI: 10.1128/JVI.00035-20
Priming of Antiviral CD8 T Cells without Effector Function by a Persistently Replicating Hepatitis C-Like Virus
Abstract
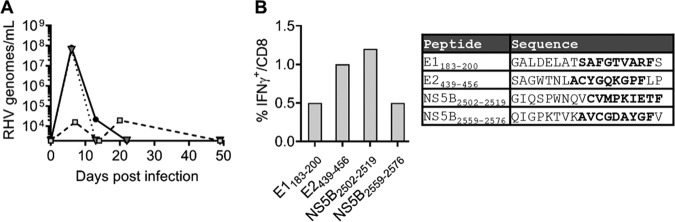
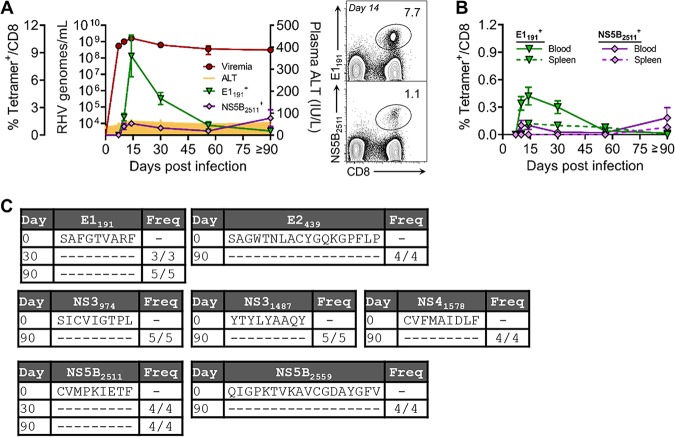
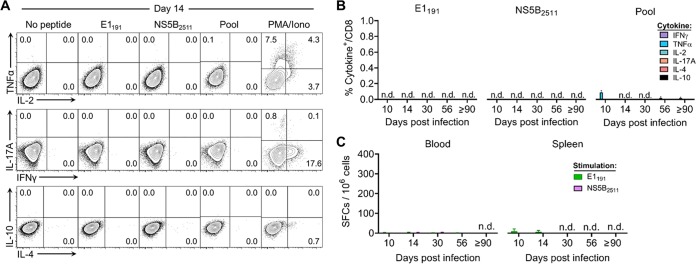
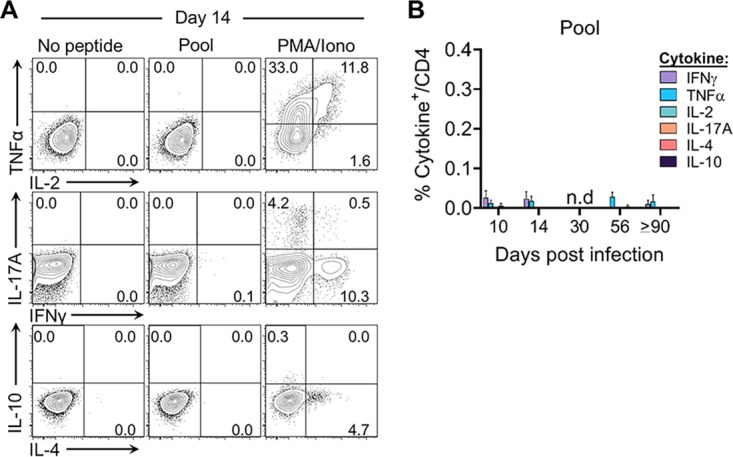
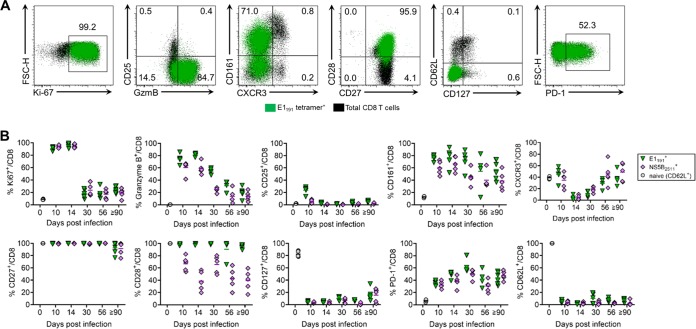
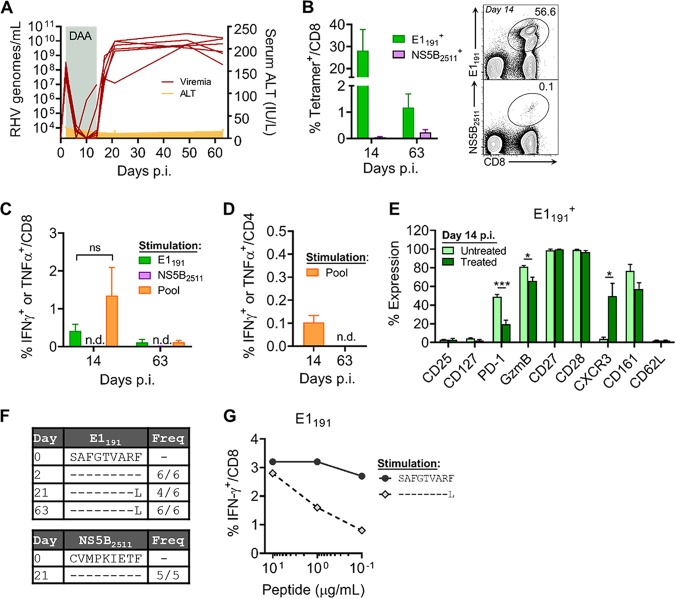
Immune-competent animal models for the hepatitis C virus (HCV) are nonexistent, impeding studies of host-virus interactions and vaccine development. Experimental infection of laboratory rats with a rodent hepacivirus isolated from Rattus norvegicus (RHV) is a promising surrogate model due to its recapitulation of HCV-like chronicity. However, several aspects of rat RHV infection remain unclear, for instance, how RHV evades host adaptive immunity to establish persistent infection. Here, we analyzed the induction, differentiation, and functionality of RHV-specific CD8 T cell responses that are essential for protection against viral persistence. Virus-specific CD8 T cells targeting dominant and subdominant major histocompatibility complex class I epitopes proliferated considerably in liver after RHV infection. These populations endured long term yet never acquired antiviral effector functions or selected for viral escape mutations. This was accompanied by the persistent upregulation of programmed cell death-1 and absent memory cell formation, consistent with a dysfunctional phenotype. Remarkably, transient suppression of RHV viremia with a direct-acting antiviral led to the priming of CD8 T cells with partial effector function, driving the selection of a viral escape variant. These data demonstrate an intrinsic abnormality within CD8 T cells primed by rat RHV infection, an effect that is governed at least partially by the magnitude of early virus replication. Thus, this model could be useful in investigating mechanisms of CD8 T cell subversion, leading to the persistence of hepatotropic pathogens such as HCV.IMPORTANCE Development of vaccines against hepatitis C virus (HCV), a major cause of cirrhosis and cancer, has been stymied by a lack of animal models. The recent discovery of an HCV-like rodent hepacivirus (RHV) enabled the development of such a model in rats. This platform recapitulates HCV hepatotropism and viral chronicity necessary for vaccine testing. Currently, there are few descriptions of RHV-specific responses and why they fail to prevent persistent infection in this model. Here, we show that RHV-specific CD8 T cells, while induced early at high magnitude, do not develop into functional effectors capable of controlling virus. This defect was partially alleviated by short-term treatment with an HCV antiviral. Thus, like HCV, RHV triggers dysfunction of virus-specific CD8 T cells that are vital for infection resolution. Additional study of this evasion strategy and how to mitigate it could enhance our understanding of hepatotropic viral infections and lead to improved vaccines and therapeutics.
Keywords: T cells; animal models; antiviral agents; antivirals; hepacivirus; hepatitis C virus; vaccines.
Copyright © 2020 American Society for Microbiology.
Figures
Similar articles
-
Adenovirus-vectored T cell vaccine for hepacivirus shows reduced effectiveness against a CD8 T cell escape variant in rats.PLoS Pathog. 2021 Mar 18;17(3):e1009391. doi: 10.1371/journal.ppat.1009391. eCollection 2021 Mar. PLoS Pathog. 2021. PMID: 33735321 Free PMC article.
-
Replicons of a Rodent Hepatitis C Model Virus Permit Selection of Highly Permissive Cells.J Virol. 2019 Sep 12;93(19):e00733-19. doi: 10.1128/JVI.00733-19. Print 2019 Oct 1. J Virol. 2019. PMID: 31292246 Free PMC article.
-
Viral persistence, liver disease, and host response in a hepatitis C-like virus rat model.Hepatology. 2018 Aug;68(2):435-448. doi: 10.1002/hep.29494. Epub 2018 May 21. Hepatology. 2018. PMID: 28859226 Free PMC article.
-
CD8+ T-Cell Responses in Hepatitis B and C: The (HLA-) A, B, and C of Hepatitis B and C.Dig Dis. 2016;34(4):396-409. doi: 10.1159/000444555. Epub 2016 May 11. Dig Dis. 2016. PMID: 27170395 Review.
-
Innate and adaptive immune responses in HCV infections.J Hepatol. 2014 Nov;61(1 Suppl):S14-25. doi: 10.1016/j.jhep.2014.06.035. Epub 2014 Nov 3. J Hepatol. 2014. PMID: 25443342 Review.
Cited by
-
Neutralization and receptor use of infectious culture-derived rat hepacivirus as a model for HCV.Hepatology. 2022 Nov;76(5):1506-1519. doi: 10.1002/hep.32535. Epub 2022 May 12. Hepatology. 2022. PMID: 35445423 Free PMC article.
-
Adenovirus-vectored T cell vaccine for hepacivirus shows reduced effectiveness against a CD8 T cell escape variant in rats.PLoS Pathog. 2021 Mar 18;17(3):e1009391. doi: 10.1371/journal.ppat.1009391. eCollection 2021 Mar. PLoS Pathog. 2021. PMID: 33735321 Free PMC article.
-
Phenotype and fate of liver-resident CD8 T cells during acute and chronic hepacivirus infection.PLoS Pathog. 2023 Oct 9;19(10):e1011697. doi: 10.1371/journal.ppat.1011697. eCollection 2023 Oct. PLoS Pathog. 2023. PMID: 37812637 Free PMC article.
-
Animal Models Used in Hepatitis C Virus Research.Int J Mol Sci. 2020 May 29;21(11):3869. doi: 10.3390/ijms21113869. Int J Mol Sci. 2020. PMID: 32485887 Free PMC article. Review.
-
Hepatitis C Virus Vaccine Research: Time to Put Up or Shut Up.Viruses. 2021 Aug 12;13(8):1596. doi: 10.3390/v13081596. Viruses. 2021. PMID: 34452460 Free PMC article. Review.
References
-
- WHO. 2017. Global hepatitis report 2017. World Health Organization, Geneva, Switzerland.
-
- Bartenschlager R, Baumert TF, Bukh J, Houghton M, Lemon SM, Lindenbach BD, Lohmann V, Moradpour D, Pietschmann T, Rice CM, Thimme R, Wakita T. 2018. Critical challenges and emerging opportunities in hepatitis C virus research in an era of potent antiviral therapy: considerations for scientists and funding agencies. Virus Res 248:53–62. doi:10.1016/j.virusres.2018.02.016. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
