

A Bivalent, Spherical Virus-Like Particle Vaccine Enhances Breadth of Immune Responses against Pathogenic Ebola Viruses in Rhesus Macaques
- PMID: 32075939
- PMCID: PMC7163155
- DOI: 10.1128/JVI.01884-19
A Bivalent, Spherical Virus-Like Particle Vaccine Enhances Breadth of Immune Responses against Pathogenic Ebola Viruses in Rhesus Macaques
Abstract
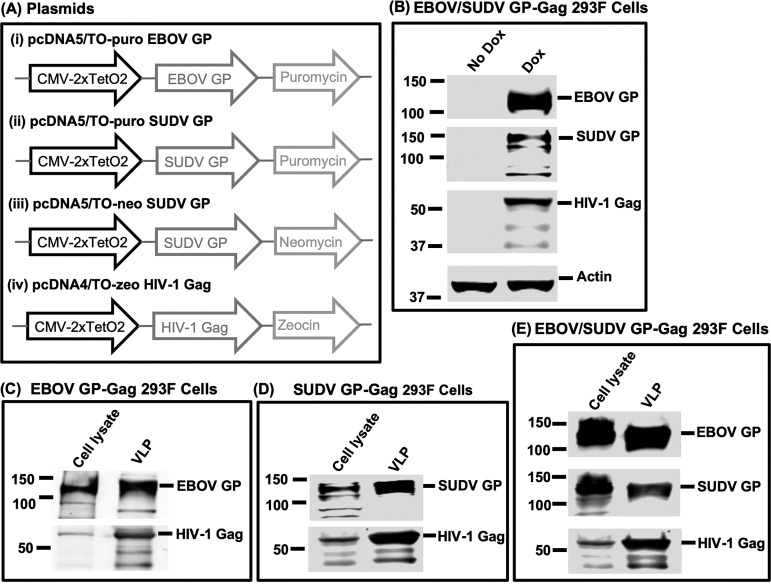
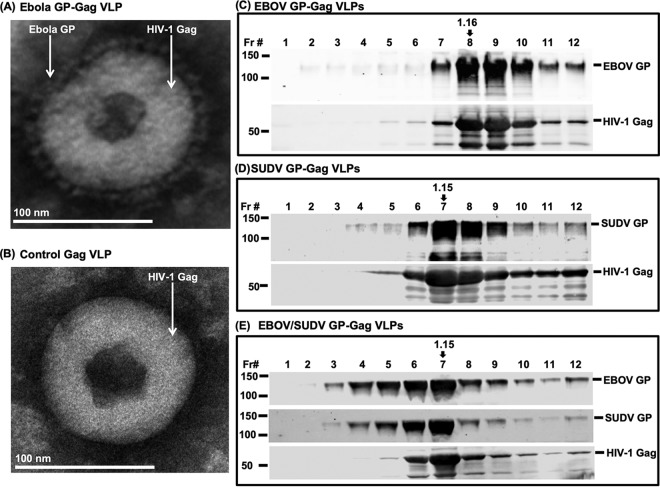
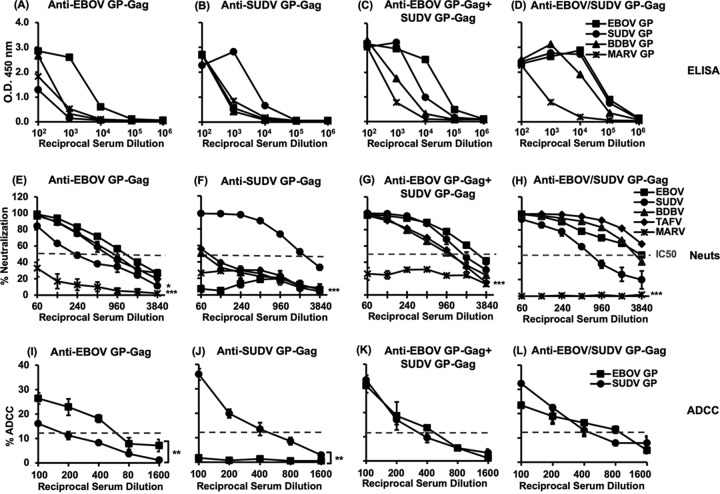
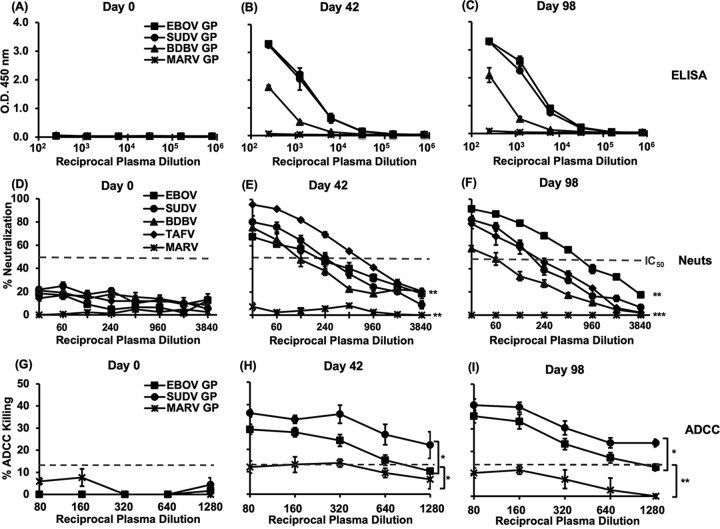
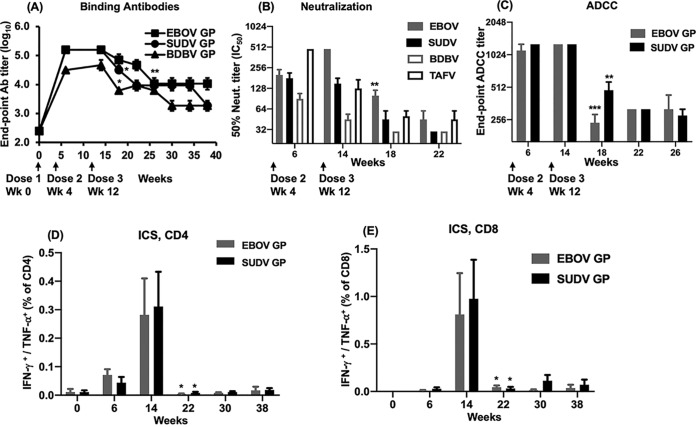
The 2013-2016 Ebola outbreak in West Africa led to accelerated efforts to develop vaccines against these highly virulent viruses. A live, recombinant vesicular stomatitis virus-based vaccine has been deployed in outbreak settings and appears highly effective. Vaccines based on replication-deficient adenovirus vectors either alone or in combination with a multivalent modified vaccinia Ankara (MVA) Ebola vaccine also appear promising and are progressing in clinical evaluation. However, the ability of current live vector-based approaches to protect against multiple pathogenic species of Ebola is not yet established, and eliciting durable responses may require additional booster vaccinations. Here, we report the development of a bivalent, spherical Ebola virus-like particle (VLP) vaccine that incorporates glycoproteins (GPs) from Zaire Ebola virus (EBOV) and Sudan Ebola virus (SUDV) and is designed to extend the breadth of immunity beyond EBOV. Immunization of rabbits with bivalent Ebola VLPs produced antibodies that neutralized all four pathogenic species of Ebola viruses and elicited antibody-dependent cell-mediated cytotoxicity (ADCC) responses against EBOV and SUDV. Vaccination of rhesus macaques with bivalent VLPs generated strong humoral immune responses, including high titers of binding, as well as neutralizing antibodies and ADCC responses. VLP vaccination led to a significant increase in the frequency of Ebola GP-specific CD4 and CD8 T cell responses. These results demonstrate that a novel bivalent Ebola VLP vaccine elicits strong humoral and cellular immune responses against pathogenic Ebola viruses and support further evaluation of this approach as a potential addition to Ebola vaccine development efforts.IMPORTANCE Ebola outbreaks result in significant morbidity and mortality in affected countries. Although several leading candidate Ebola vaccines have been developed and advanced in clinical testing, additional vaccine candidates may be needed to provide protection against different Ebola species and to extend the durability of protection. A novel approach demonstrated here is to express two genetically diverse glycoproteins on a spherical core, generating a vaccine that can broaden immune responses against known pathogenic Ebola viruses. This approach provides a new method to broaden and potentially extend protective immune responses against Ebola viruses.
Keywords: Bundibugyo virus; Ebola glycoprotein; Ebola vaccine; Ebola virus; Ebola virus disease; Marburg virus; Sudan virus; Tai Forest virus; virus-like particles.
Copyright © 2020 American Society for Microbiology.
Figures
Similar articles
-
Distinct Immunogenicity and Efficacy of Poxvirus-Based Vaccine Candidates against Ebola Virus Expressing GP and VP40 Proteins.J Virol. 2018 May 14;92(11):e00363-18. doi: 10.1128/JVI.00363-18. Print 2018 Jun 1. J Virol. 2018. PMID: 29514907 Free PMC article.
-
Recombinant Modified Vaccinia Virus Ankara Generating Ebola Virus-Like Particles.J Virol. 2017 May 12;91(11):e00343-17. doi: 10.1128/JVI.00343-17. Print 2017 Jun 1. J Virol. 2017. PMID: 28331098 Free PMC article.
-
Antibody-Mediated Protective Mechanisms Induced by a Trivalent Parainfluenza Virus-Vectored Ebolavirus Vaccine.J Virol. 2019 Feb 5;93(4):e01845-18. doi: 10.1128/JVI.01845-18. Print 2019 Feb 15. J Virol. 2019. PMID: 30518655 Free PMC article.
-
Correlates of vaccine-induced protective immunity against Ebola virus disease.Semin Immunol. 2018 Oct;39:65-72. doi: 10.1016/j.smim.2018.07.003. Epub 2018 Jul 21. Semin Immunol. 2018. PMID: 30041831 Review.
-
Recent advances in the development of vaccines for Ebola virus disease.Virus Res. 2016 Jan 4;211:174-85. doi: 10.1016/j.virusres.2015.10.021. Epub 2015 Oct 24. Virus Res. 2016. PMID: 26596227 Review.
Cited by
-
Viral vectored vaccines: design, development, preventive and therapeutic applications in human diseases.Signal Transduct Target Ther. 2023 Apr 7;8(1):149. doi: 10.1038/s41392-023-01408-5. Signal Transduct Target Ther. 2023. PMID: 37029123 Free PMC article. Review.
-
Human Vaccines & Immunotherapeutics: news.Hum Vaccin Immunother. 2020 Jun 2;16(6):1226-1227. doi: 10.1080/21645515.2020.1778422. Hum Vaccin Immunother. 2020. PMID: 32598246 Free PMC article. No abstract available.
-
Innate Immune Sensing of Viruses and Its Consequences for the Central Nervous System.Viruses. 2021 Jan 23;13(2):170. doi: 10.3390/v13020170. Viruses. 2021. PMID: 33498715 Free PMC article. Review.
-
Nanoparticles and Antiviral Vaccines.Vaccines (Basel). 2023 Dec 27;12(1):30. doi: 10.3390/vaccines12010030. Vaccines (Basel). 2023. PMID: 38250843 Free PMC article. Review.
-
The development and kinetics of functional antibody-dependent cell-mediated cytotoxicity (ADCC) to SARS-CoV-2 spike protein.Virology. 2021 Jul;559:1-9. doi: 10.1016/j.virol.2021.03.009. Epub 2021 Mar 19. Virology. 2021. PMID: 33774551 Free PMC article.
References
-
- WHO Ebola Response Team, Agua-Agum J, Allegranzi B, Ariyarajah A, Aylward R, Blake IM, Barboza P, Bausch D, Brennan RJ, Clement P, Coffey P, Cori A, Donnelly CA, Dorigatti I, Drury P, Durski K, Dye C, Eckmanns T, Ferguson NM, Fraser C, Garcia E, Garske T, Gasasira A, Gurry C, Hamblion E, Hinsley W, Holden R, Holmes D, Hugonnet S, Jaramillo Gutierrez G, Jombart T, Kelley E, Santhana R, Mahmoud N, Mills HL, Mohamed Y, Musa E, Naidoo D, Nedjati-Gilani G, Newton E, Norton I, Nouvellet P, Perkins D, Perkins M, Riley S, Schumacher D, Shah A, Tang M, Varsaneux O, Van Kerkhove MD. 2016. After Ebola in West Africa–unpredictable risks, preventable epidemics. N Engl J Med 375:587–596. doi:10.1056/NEJMsr1513109. - DOI - PubMed
-
- Henao-Restrepo AM, Camacho A, Longini IM, Watson CH, Edmunds WJ, Egger M, Carroll MW, Dean NE, Diatta I, Doumbia M, Draguez B, Duraffour S, Enwere G, Grais R, Gunther S, Gsell P-S, Hossmann S, Watle SV, Kondé MK, Kéïta S, Kone S, Kuisma E, Levine MM, Mandal S, Mauget T, Norheim G, Riveros X, Soumah A, Trelle S, Vicari AS, Røttingen J-A, Kieny M-P. 2017. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ca Suffit!). Lancet 389:505–518. doi:10.1016/S0140-6736(16)32621-6. - DOI - PMC - PubMed
-
- Ledgerwood JE, VRC 207 Study Team, DeZure AD, Stanley DA, Coates EE, Novik L, Enama ME, Berkowitz NM, Hu Z, Joshi G, Ploquin A, Sitar S, Gordon IJ, Plummer SA, Holman LA, Hendel CS, Yamshchikov G, Roman F, Nicosia A, Colloca S, Cortese R, Bailer RT, Schwartz RM, Roederer M, Mascola JR, Koup RA, Sullivan NJ, Graham BS, Team V. 2017. Chimpanzee adenovirus vector ebola vaccine. N Engl J Med 376:928–938. doi:10.1056/NEJMoa1410863. - DOI - PubMed
-
- Li JX, Hou LH, Meng FY, Wu SP, Hu YM, Liang Q, Chu K, Zhang Z, Xu JJ, Tang R, Wang WJ, Liu P, Hu JL, Luo L, Jiang R, Zhu FC, Chen W. 2017. Immunity duration of a recombinant adenovirus type-5 vector-based Ebola vaccine and a homologous prime-boost immunisation in healthy adults in China: final report of a randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Glob Health 5:e324–e334. doi:10.1016/S2214-109X(16)30367-9. - DOI - PubMed
-
- Milligan ID, Gibani MM, Sewell R, Clutterbuck EA, Campbell D, Plested E, Nuthall E, Voysey M, Silva-Reyes L, McElrath MJ, De Rosa SC, Frahm N, Cohen KW, Shukarev G, Orzabal N, van Duijnhoven W, Truyers C, Bachmayer N, Splinter D, Samy N, Pau MG, Schuitemaker H, Luhn K, Callendret B, Van Hoof J, Douoguih M, Ewer K, Angus B, Pollard AJ, Snape MD. 2016. Safety and immunogenicity of novel adenovirus type 26- and modified vaccinia Ankara-vectored Ebola vaccines: a randomized clinical trial. JAMA 315:1610–1623. doi:10.1001/jama.2016.4218. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
