

Current Status of Latency Reversing Agents Facing the Heterogeneity of HIV-1 Cellular and Tissue Reservoirs
- PMID: 32038533
- PMCID: PMC6993040
- DOI: 10.3389/fmicb.2019.03060
Current Status of Latency Reversing Agents Facing the Heterogeneity of HIV-1 Cellular and Tissue Reservoirs
Abstract
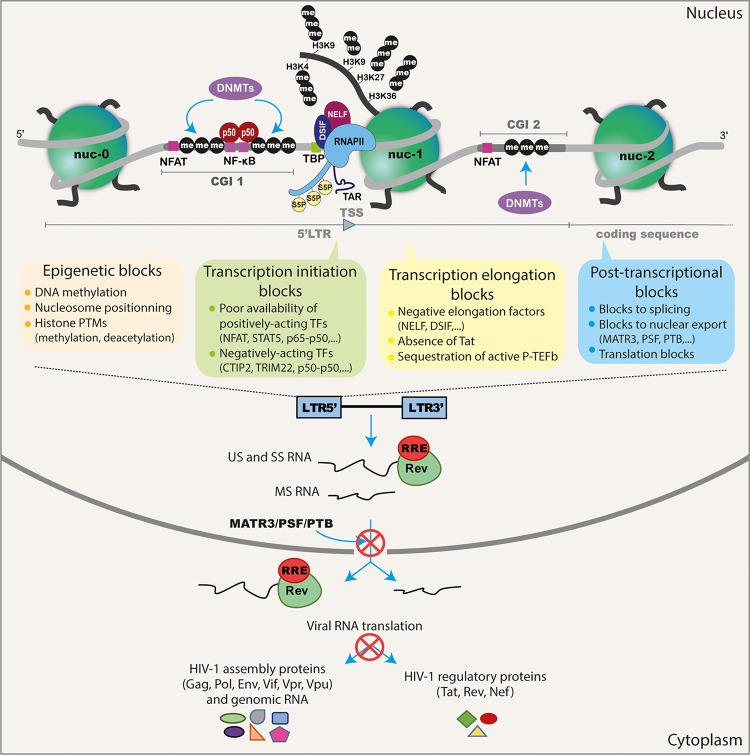
One of the most explored therapeutic approaches aimed at eradicating HIV-1 reservoirs is the "shock and kill" strategy which is based on HIV-1 reactivation in latently-infected cells ("shock" phase) while maintaining antiretroviral therapy (ART) in order to prevent spreading of the infection by the neosynthesized virus. This kind of strategy allows for the "kill" phase, during which latently-infected cells die from viral cytopathic effects or from host cytolytic effector mechanisms following viral reactivation. Several latency reversing agents (LRAs) with distinct mechanistic classes have been characterized to reactivate HIV-1 viral gene expression. Some LRAs have been tested in terms of their potential to purge latent HIV-1 in vivo in clinical trials, showing that reversing HIV-1 latency is possible. However, LRAs alone have failed to reduce the size of the viral reservoirs. Together with the inability of the immune system to clear the LRA-activated reservoirs and the lack of specificity of these LRAs, the heterogeneity of the reservoirs largely contributes to the limited success of clinical trials using LRAs. Indeed, HIV-1 latency is established in numerous cell types that are characterized by distinct phenotypes and metabolic properties, and these are influenced by patient history. Hence, the silencing mechanisms of HIV-1 gene expression in these cellular and tissue reservoirs need to be better understood to rationally improve this cure strategy and hopefully reach clinical success.
Keywords: HIV-1; cure; heterogeneity; latency; latency reversing agents; reservoirs.
Copyright © 2020 Ait-Ammar, Kula, Darcis, Verdikt, De Wit, Gautier, Mallon, Marcello, Rohr and Van Lint.
Figures
Similar articles
-
Posttranscriptional Regulation of HIV-1 Gene Expression during Replication and Reactivation from Latency by Nuclear Matrix Protein MATR3.mBio. 2018 Nov 13;9(6):e02158-18. doi: 10.1128/mBio.02158-18. mBio. 2018. PMID: 30425153 Free PMC article.
-
The Current Status of Latency Reversing Agents for HIV-1 Remission.Annu Rev Virol. 2021 Sep 29;8(1):491-514. doi: 10.1146/annurev-virology-091919-103029. Annu Rev Virol. 2021. PMID: 34586875
-
Alternate NF-κB-Independent Signaling Reactivation of Latent HIV-1 Provirus.J Virol. 2019 Aug 28;93(18):e00495-19. doi: 10.1128/JVI.00495-19. Print 2019 Sep 15. J Virol. 2019. PMID: 31243131 Free PMC article.
-
Fighting HIV-1 Persistence: At the Crossroads of "Shoc-K and B-Lock".Pathogens. 2021 Nov 20;10(11):1517. doi: 10.3390/pathogens10111517. Pathogens. 2021. PMID: 34832672 Free PMC article. Review.
-
Shocking HIV-1 with immunomodulatory latency reversing agents.Semin Immunol. 2021 Jan;51:101478. doi: 10.1016/j.smim.2021.101478. Epub 2021 May 8. Semin Immunol. 2021. PMID: 33972164 Review.
Cited by
-
Small molecule inhibitors of transcriptional cyclin-dependent kinases impose HIV-1 latency, presenting "block and lock" treatment strategies.Antimicrob Agents Chemother. 2024 Mar 6;68(3):e0107223. doi: 10.1128/aac.01072-23. Epub 2024 Feb 6. Antimicrob Agents Chemother. 2024. PMID: 38319085 Free PMC article.
-
Elevation of cell-associated HIV-1 transcripts in CSF CD4+ T cells, despite effective antiretroviral therapy, is linked to brain injury.Proc Natl Acad Sci U S A. 2022 Nov 29;119(48):e2210584119. doi: 10.1073/pnas.2210584119. Epub 2022 Nov 21. Proc Natl Acad Sci U S A. 2022. PMID: 36413502 Free PMC article.
-
Aminobisphosphonates reactivate the latent reservoir in people living with HIV-1.bioRxiv [Preprint]. 2023 Feb 7:2023.02.07.527421. doi: 10.1101/2023.02.07.527421. bioRxiv. 2023. Update in: Front Immunol. 2023 Sep 06;14:1219250. doi: 10.3389/fimmu.2023.1219250 PMID: 36798291 Free PMC article. Updated. Preprint.
-
Role of TLRs in HIV-1 Infection and Potential of TLR Agonists in HIV-1 Vaccine Development and Treatment Strategies.Pathogens. 2023 Jan 5;12(1):92. doi: 10.3390/pathogens12010092. Pathogens. 2023. PMID: 36678440 Free PMC article. Review.
-
Enhanced Cross-Reactive and Polyfunctional Effector-Memory T Cell Responses by ICVAX-a Human PD1-Based Bivalent HIV-1 Gag-p41 Mosaic DNA Vaccine.J Virol. 2022 Apr 13;96(7):e0216121. doi: 10.1128/jvi.02161-21. Epub 2022 Mar 17. J Virol. 2022. PMID: 35297660 Free PMC article.
References
-
- Abdel-Mohsen M., Kuri-Cervantes L., Grau-Exposito J., Spivak A. M., Nell R. A., Tomescu C., et al. (2018). CD32 is expressed on cells with transcriptionally active HIV but does not enrich for HIV DNA in resting T cells. Sci. Transl. Med. 10:eaar6759. 10.1126/scitranslmed.aar6759 - DOI - PMC - PubMed
-
- Adams M., Sharmeen L., Kimpton J., Romeo J. M., Garcia J. V., Peterlin B. M., et al. (1994). Cellular latency in human immunodeficiency virus-infected individuals with high CD4 levels can be detected by the presence of promoter-proximal transcripts. Proc. Natl. Acad. Sci. U.S.A. 91 3862–3866. 10.1073/pnas.91.9.3862 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources