

Selective reconstitution of IFN‑γ gene function in Ncr1+ NK cells is sufficient to control systemic vaccinia virus infection
- PMID: 32023327
- PMCID: PMC7028289
- DOI: 10.1371/journal.ppat.1008279
Selective reconstitution of IFN‑γ gene function in Ncr1+ NK cells is sufficient to control systemic vaccinia virus infection
Abstract
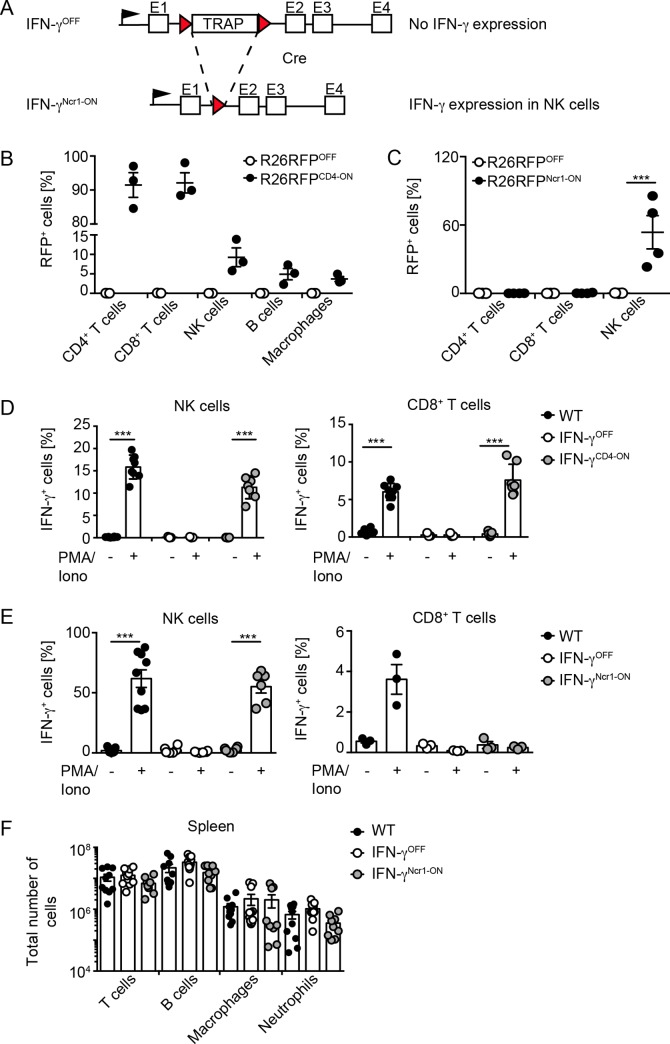
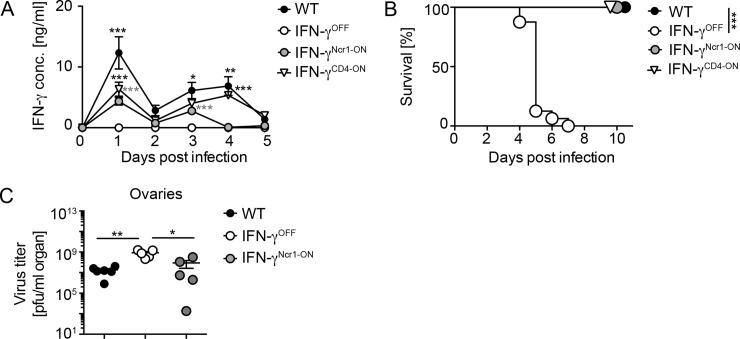
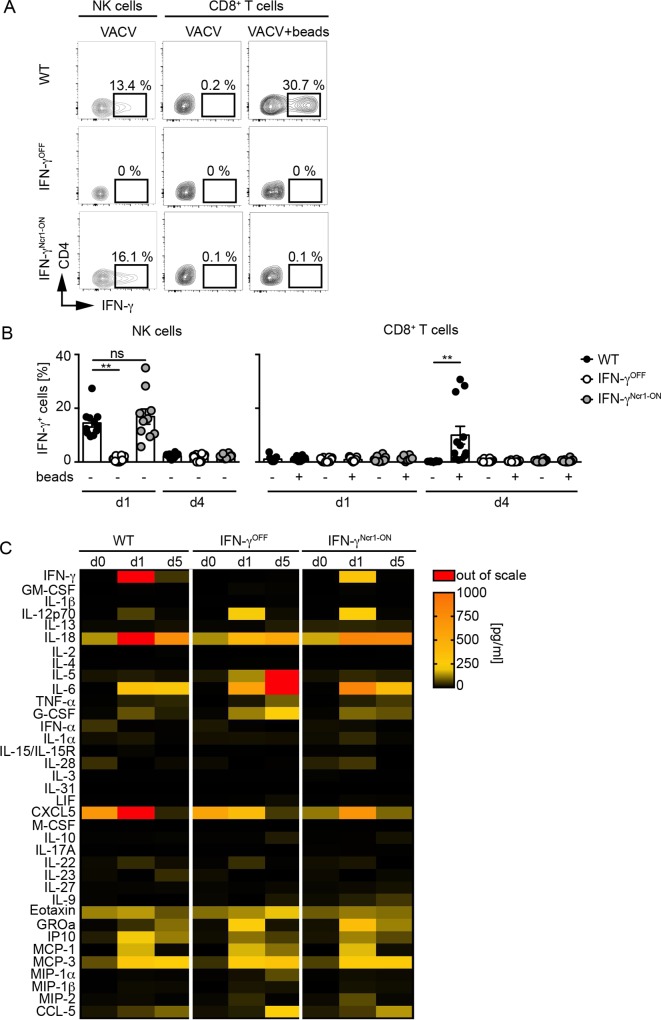
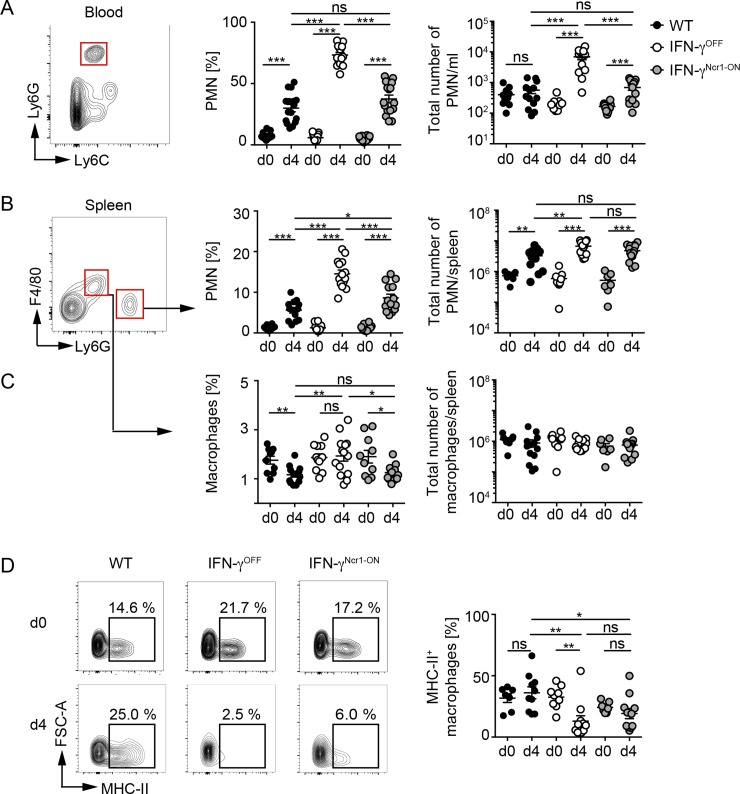
IFN-γ is an enigmatic cytokine that shows direct anti-viral effects, confers upregulation of MHC-II and other components relevant for antigen presentation, and that adjusts the composition and balance of complex cytokine responses. It is produced during immune responses by innate as well as adaptive immune cells and can critically affect the course and outcome of infectious diseases, autoimmunity, and cancer. To selectively analyze the function of innate immune cell-derived IFN-γ, we generated conditional IFN-γOFF mice, in which endogenous IFN-γ expression is disrupted by a loxP flanked gene trap cassette inserted into the first intron of the IFN-γ gene. IFN-γOFF mice were intercrossed with Ncr1-Cre or CD4-Cre mice that express Cre mainly in NK cells (IFN-γNcr1-ON mice) or T cells (IFN-γCD4-ON mice), respectively. Rosa26RFP reporter mice intercrossed with Ncr1-Cre mice showed selective RFP expression in more than 80% of the NK cells, while upon intercrossing with CD4-Cre mice abundant RFP expression was detected in T cells, but also to a minor extent in other immune cell subsets. Previous studies showed that IFN-γ expression is needed to promote survival of vaccinia virus (VACV) infection. Interestingly, during VACV infection of wild type and IFN-γCD4-ON mice two waves of serum IFN-γ were induced that peaked on day 1 and day 3/4 after infection. Similarly, VACV infected IFN-γNcr1-ON mice mounted two waves of IFN-γ responses, of which the first one was moderately and the second one profoundly reduced when compared with WT mice. Furthermore, IFN-γNcr1-ON as well as IFN-γCD4-ON mice survived VACV infection, whereas IFN-γOFF mice did not. As expected, ex vivo analysis of splenocytes derived from VACV infected IFN-γNcr1-ON mice showed IFN-γ expression in NK cells, but not T cells, whereas IFN-γOFF mice showed IFN-γ expression neither in NK cells nor T cells. VACV infected IFN-γNcr1-ON mice mounted normal cytokine responses, restored neutrophil accumulation, and showed normal myeloid cell distribution in blood and spleen. Additionally, in these mice normal MHC-II expression was detected on peripheral macrophages, whereas IFN-γOFF mice did not show MHC-II expression on such cells. In conclusion, upon VACV infection Ncr1 positive cells including NK cells mount two waves of early IFN-γ responses that are sufficient to promote the induction of protective anti-viral immunity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Natural Killer Cells and Innate Interferon Gamma Participate in the Host Defense against Respiratory Vaccinia Virus Infection.J Virol. 2015 Oct 14;90(1):129-41. doi: 10.1128/JVI.01894-15. Print 2016 Jan 1. J Virol. 2015. PMID: 26468539 Free PMC article.
-
Insufficient Innate Immunity Contributes to the Susceptibility of the Castaneous Mouse to Orthopoxvirus Infection.J Virol. 2017 Sep 12;91(19):e01042-17. doi: 10.1128/JVI.01042-17. Print 2017 Oct 1. J Virol. 2017. PMID: 28747505 Free PMC article.
-
Vaccinia virus decreases major histocompatibility complex (MHC) class II antigen presentation, T-cell priming, and peptide association with MHC class II.Immunology. 2009 Nov;128(3):381-92. doi: 10.1111/j.1365-2567.2009.03120.x. Immunology. 2009. PMID: 20067538 Free PMC article.
-
Interferon-gamma: biologic functions and HCV therapy (type I/II) (1 of 2 parts).Clin Ter. 2006 Jul-Aug;157(4):377-86. Clin Ter. 2006. Retraction in: Clin Ter. 2008 May-Jun;159(3):207. PMID: 17051976 Retracted. Review.
-
Definition of epitopes and antigens recognized by vaccinia specific immune responses: their conservation in variola virus sequences, and use as a model system to study complex pathogens.Vaccine. 2009 Dec 30;27 Suppl 6(Suppl 6):G21-6. doi: 10.1016/j.vaccine.2009.10.011. Vaccine. 2009. PMID: 20006135 Free PMC article. Review.
Cited by
-
Antiviral Activities of Group I Innate Lymphoid Cells.J Mol Biol. 2022 Mar 30;434(6):167266. doi: 10.1016/j.jmb.2021.167266. Epub 2021 Sep 22. J Mol Biol. 2022. PMID: 34562465 Free PMC article. Review.
-
Forms and Methods for Interferon's Encapsulation.Pharmaceutics. 2021 Sep 22;13(10):1533. doi: 10.3390/pharmaceutics13101533. Pharmaceutics. 2021. PMID: 34683824 Free PMC article. Review.
-
IL-15 Priming Alters IFN-γ Regulation in Murine NK Cells.J Immunol. 2023 Nov 15;211(10):1481-1493. doi: 10.4049/jimmunol.2300283. J Immunol. 2023. PMID: 37747317 Free PMC article.
-
Autologous CIK cells combined with chemotherapy as the first-line treatment for locally advanced or metastatic gastric cancer is safe and feasible.Front Immunol. 2023 Nov 1;14:1267369. doi: 10.3389/fimmu.2023.1267369. eCollection 2023. Front Immunol. 2023. PMID: 38022664 Free PMC article. Clinical Trial.
-
Segmented filamentous bacteria-induced epithelial MHCII regulates cognate CD4+ IELs and epithelial turnover.J Exp Med. 2024 Jan 1;221(1):e20230194. doi: 10.1084/jem.20230194. Epub 2023 Oct 30. J Exp Med. 2024. PMID: 37902602 Free PMC article.
References
-
- Gajewski TF, Fitch FW. Anti-proliferative effect of IFN-gamma in immune regulation. I. IFN-gamma inhibits the proliferation of Th2 but not Th1 murine helper T lymphocyte clones. J Immunol. 1988;140(12):4245–52. Epub 1988/06/15. . - PubMed
-
- Orange JS, Wang B, Terhorst C, Biron CA. Requirement for natural killer cell-produced interferon gamma in defense against murine cytomegalovirus infection and enhancement of this defense pathway by interleukin 12 administration. J Exp Med. 1995;182(4):1045–56. Epub 1995/10/01. 10.1084/jem.182.4.1045 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
