

Coral microbiome composition along the northern Red Sea suggests high plasticity of bacterial and specificity of endosymbiotic dinoflagellate communities
- PMID: 32008576
- PMCID: PMC6996193
- DOI: 10.1186/s40168-019-0776-5
Coral microbiome composition along the northern Red Sea suggests high plasticity of bacterial and specificity of endosymbiotic dinoflagellate communities
Erratum in
-
Correction to: Coral microbiome composition along the northern Red Sea suggests high plasticity of bacterial and specificity of endosymbiotic dinoflagellate communities.Microbiome. 2020 Feb 21;8(1):24. doi: 10.1186/s40168-020-00807-y. Microbiome. 2020. PMID: 32085815 Free PMC article.
Abstract
Background: The capacity of reef-building corals to tolerate (or adapt to) heat stress is a key factor determining their resilience to future climate change. Changes in coral microbiome composition (particularly for microalgal endosymbionts and bacteria) is a potential mechanism that may assist corals to thrive in warm waters. The northern Red Sea experiences extreme temperatures anomalies, yet corals in this area rarely bleach suggesting possible refugia to climate change. However, the coral microbiome composition, and how it relates to the capacity to thrive in warm waters in this region, is entirely unknown.
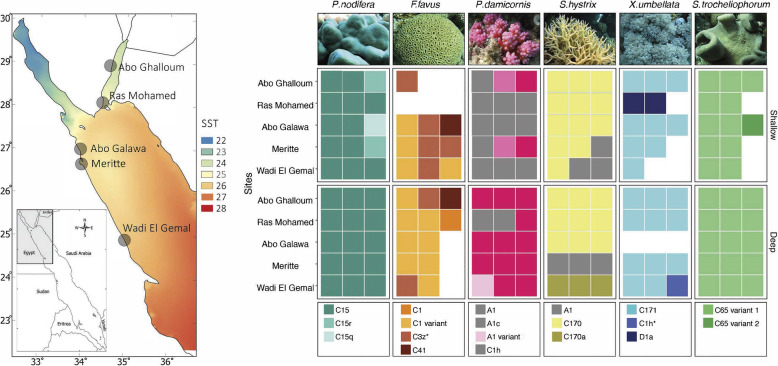
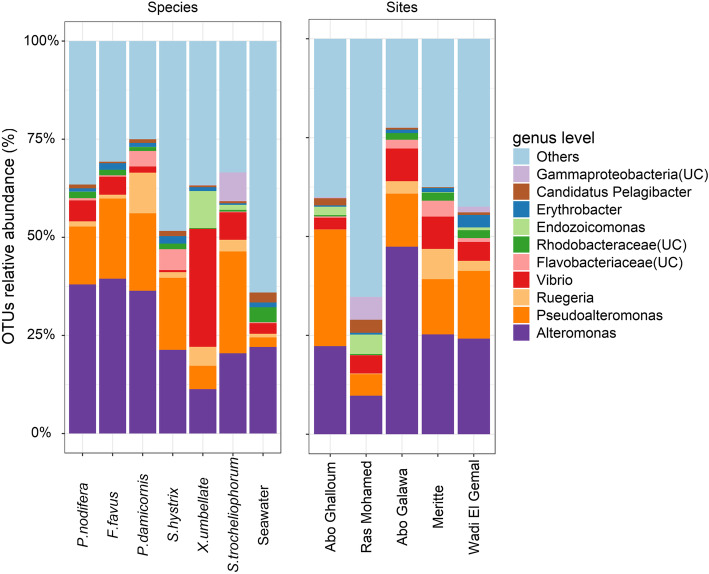
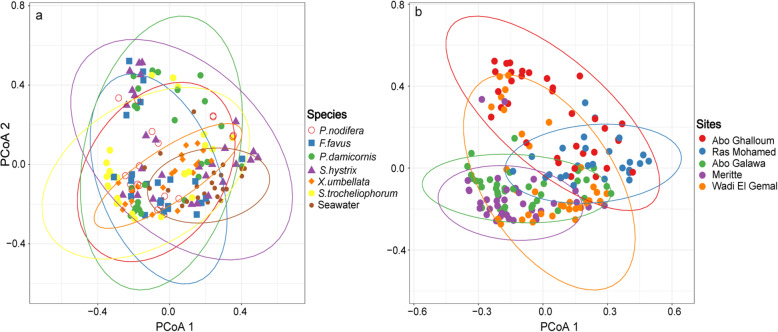
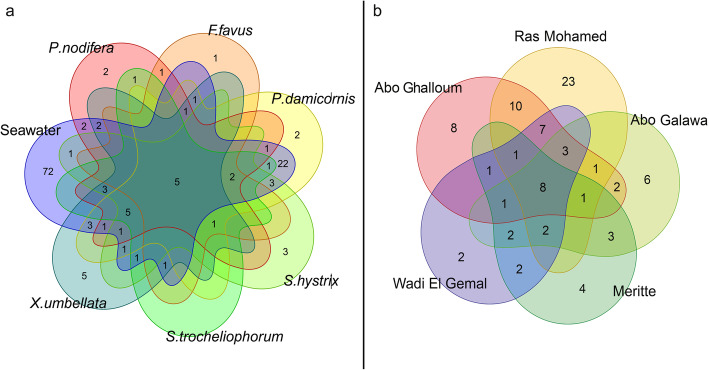
Results: We investigated microbiomes for six coral species (Porites nodifera, Favia favus, Pocillopora damicornis, Seriatopora hystrix, Xenia umbellata, and Sarcophyton trocheliophorum) from five sites in the northern Red Sea spanning 4° of latitude and summer mean temperature ranges from 26.6 °C to 29.3 °C. A total of 19 distinct dinoflagellate endosymbionts were identified as belonging to three genera in the family Symbiodiniaceae (Symbiodinium, Cladocopium, and Durusdinium). Of these, 86% belonged to the genus Cladocopium, with notably five novel types (19%). The endosymbiont community showed a high degree of host-specificity despite the latitudinal gradient. In contrast, the diversity and composition of bacterial communities of the surface mucus layer (SML)-a compartment particularly sensitive to environmental change-varied significantly between sites, however for any given coral was species-specific.
Conclusion: The conserved endosymbiotic community suggests high physiological plasticity to support holobiont productivity across the different latitudinal regimes. Further, the presence of five novel algal endosymbionts suggests selection of certain genotypes (or genetic adaptation) within the semi-isolated Red Sea. In contrast, the dynamic composition of bacteria associated with the SML across sites may contribute to holobiont function and broaden the ecological niche. In doing so, SML bacterial communities may aid holobiont local acclimatization (or adaptation) by readily responding to changes in the host environment. Our study provides novel insight about the selective and endemic nature of coral microbiomes along the northern Red Sea refugia.
Keywords: 16S rRNA gene profiling; Climate change; Coral acclimatization; Future Oceans; Holobiont; Microbial community; Symbiodiniaceae.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Microbiome community and complexity indicate environmental gradient acclimatisation and potential microbial interaction of endemic coral holobionts in the South China Sea.Sci Total Environ. 2021 Apr 15;765:142690. doi: 10.1016/j.scitotenv.2020.142690. Epub 2020 Oct 3. Sci Total Environ. 2021. PMID: 33071127
-
Spatio-temporal analyses of Symbiodinium physiology of the coral Pocillopora verrucosa along large-scale nutrient and temperature gradients in the Red Sea.PLoS One. 2014 Aug 19;9(8):e103179. doi: 10.1371/journal.pone.0103179. eCollection 2014. PLoS One. 2014. PMID: 25137123 Free PMC article.
-
Thermal regime and host clade, rather than geography, drive Symbiodinium and bacterial assemblages in the scleractinian coral Pocillopora damicornis sensu lato.Microbiome. 2018 Feb 20;6(1):39. doi: 10.1186/s40168-018-0423-6. Microbiome. 2018. PMID: 29463295 Free PMC article.
-
Coral evolutionary responses to microbial symbioses.Philos Trans R Soc Lond B Biol Sci. 2020 Sep 28;375(1808):20190591. doi: 10.1098/rstb.2019.0591. Epub 2020 Aug 10. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32772672 Free PMC article. Review.
-
Coral-the world's most diverse symbiotic ecosystem.Mol Ecol. 2015 Nov;24(21):5330-47. doi: 10.1111/mec.13400. Epub 2015 Oct 21. Mol Ecol. 2015. PMID: 26414414 Review.
Cited by
-
Bacterial and Symbiodiniaceae communities' variation in corals with distinct traits and geographical distribution.Sci Rep. 2024 Oct 16;14(1):24319. doi: 10.1038/s41598-024-70121-2. Sci Rep. 2024. PMID: 39414857 Free PMC article.
-
Microbiome characterization of defensive tissues in the model anemone Exaiptasia diaphana.BMC Microbiol. 2021 May 21;21(1):152. doi: 10.1186/s12866-021-02211-4. BMC Microbiol. 2021. PMID: 34020587 Free PMC article.
-
Metagenomic Analysis of the Species Composition and Seasonal Distribution of Marine Dinoflagellate Communities in Four Korean Coastal Regions.Microorganisms. 2022 Jul 19;10(7):1459. doi: 10.3390/microorganisms10071459. Microorganisms. 2022. PMID: 35889179 Free PMC article.
-
Contrasting Microbiome Dynamics of Putative Denitrifying Bacteria in Two Octocoral Species Exposed to Dissolved Organic Carbon (DOC) and Warming.Appl Environ Microbiol. 2022 Jan 25;88(2):e0188621. doi: 10.1128/AEM.01886-21. Epub 2021 Nov 17. Appl Environ Microbiol. 2022. PMID: 34788073 Free PMC article.
-
Capacity of deep-sea corals to obtain nutrition from cold seeps aligned with microbiome reorganization.Glob Chang Biol. 2023 Jan;29(1):189-205. doi: 10.1111/gcb.16447. Epub 2022 Oct 21. Glob Chang Biol. 2023. PMID: 36271605 Free PMC article.
References
-
- Hughes TP, Anderson KD, Connolly SR, Heron SF, Kerry JT, Lough JM, et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science. 2018;359:80–83. - PubMed
-
- Hughes TP, Kerry JT, Álvarez-Noriega M, Álvarez-romero JG, Anderson KD, Baird A, et al. Global warming and recurrent mass bleaching of corals. Nature. 2017;543:373–377. - PubMed
-
- Dixon GB, Davies SW, Aglyamova GA, Meyer E, Bay LK, Matz MV. Coral reefs. Genomic determinants of coral heat tolerance across latitudes. Science. 2015;348:1460–1462. - PubMed
-
- Osman EO, Smith DJ, Ziegler M, Kürten B, Conrad C, El-Haddad KM, et al. Thermal refugia against coral bleaching throughout the northern Red Sea. Glob Chang Biol. 2018;24:474–484. - PubMed
-
- Oliver TA, Palumbi SR. Do fluctuating temperature environments elevate coral thermal tolerance? Coral Reefs. 2011;30:429–440.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
