

Receptor Recognition by the Novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavirus
- PMID: 31996437
- PMCID: PMC7081895
- DOI: 10.1128/JVI.00127-20
Receptor Recognition by the Novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavirus
Abstract
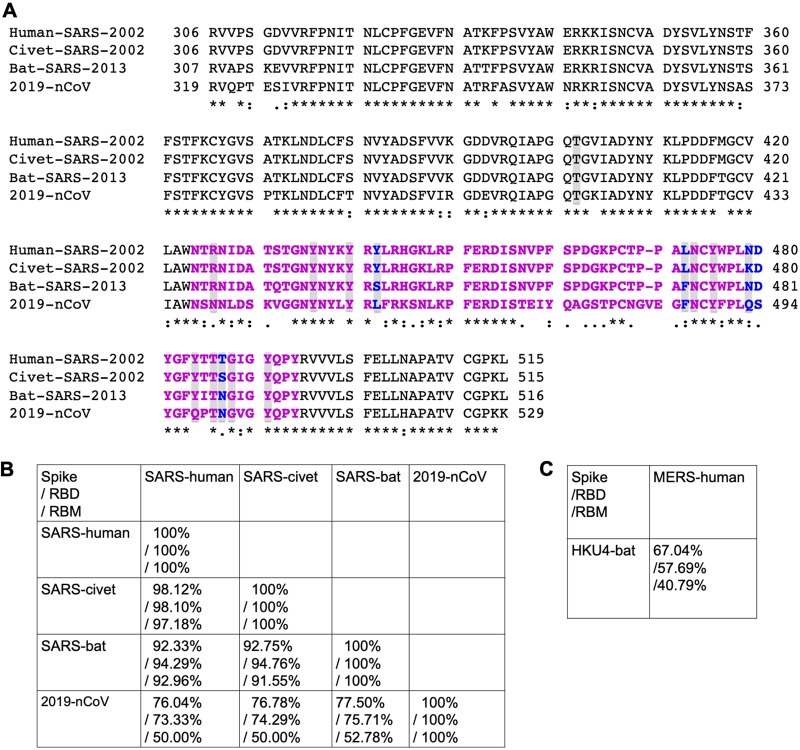
Recently, a novel coronavirus (2019-nCoV) has emerged from Wuhan, China, causing symptoms in humans similar to those caused by severe acute respiratory syndrome coronavirus (SARS-CoV). Since the SARS-CoV outbreak in 2002, extensive structural analyses have revealed key atomic-level interactions between the SARS-CoV spike protein receptor-binding domain (RBD) and its host receptor angiotensin-converting enzyme 2 (ACE2), which regulate both the cross-species and human-to-human transmissions of SARS-CoV. Here, we analyzed the potential receptor usage by 2019-nCoV, based on the rich knowledge about SARS-CoV and the newly released sequence of 2019-nCoV. First, the sequence of 2019-nCoV RBD, including its receptor-binding motif (RBM) that directly contacts ACE2, is similar to that of SARS-CoV, strongly suggesting that 2019-nCoV uses ACE2 as its receptor. Second, several critical residues in 2019-nCoV RBM (particularly Gln493) provide favorable interactions with human ACE2, consistent with 2019-nCoV's capacity for human cell infection. Third, several other critical residues in 2019-nCoV RBM (particularly Asn501) are compatible with, but not ideal for, binding human ACE2, suggesting that 2019-nCoV has acquired some capacity for human-to-human transmission. Last, while phylogenetic analysis indicates a bat origin of 2019-nCoV, 2019-nCoV also potentially recognizes ACE2 from a diversity of animal species (except mice and rats), implicating these animal species as possible intermediate hosts or animal models for 2019-nCoV infections. These analyses provide insights into the receptor usage, cell entry, host cell infectivity and animal origin of 2019-nCoV and may help epidemic surveillance and preventive measures against 2019-nCoV.IMPORTANCE The recent emergence of Wuhan coronavirus (2019-nCoV) puts the world on alert. 2019-nCoV is reminiscent of the SARS-CoV outbreak in 2002 to 2003. Our decade-long structural studies on the receptor recognition by SARS-CoV have identified key interactions between SARS-CoV spike protein and its host receptor angiotensin-converting enzyme 2 (ACE2), which regulate both the cross-species and human-to-human transmissions of SARS-CoV. One of the goals of SARS-CoV research was to build an atomic-level iterative framework of virus-receptor interactions to facilitate epidemic surveillance, predict species-specific receptor usage, and identify potential animal hosts and animal models of viruses. Based on the sequence of 2019-nCoV spike protein, we apply this predictive framework to provide novel insights into the receptor usage and likely host range of 2019-nCoV. This study provides a robust test of this reiterative framework, providing the basic, translational, and public health research communities with predictive insights that may help study and battle this novel 2019-nCoV.
Keywords: 2019-nCoV; SARS coronavirus; angiotensin-converting enzyme 2; animal reservoir; cross-species transmission; human-to-human transmission.
Copyright © 2020 American Society for Microbiology.
Figures



Similar articles
-
Composition and divergence of coronavirus spike proteins and host ACE2 receptors predict potential intermediate hosts of SARS-CoV-2.J Med Virol. 2020 Jun;92(6):595-601. doi: 10.1002/jmv.25726. Epub 2020 Mar 11. J Med Virol. 2020. PMID: 32100877 Free PMC article.
-
Comparison of Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein Binding to ACE2 Receptors from Human, Pets, Farm Animals, and Putative Intermediate Hosts.J Virol. 2020 Jul 16;94(15):e00831-20. doi: 10.1128/JVI.00831-20. Print 2020 Jul 16. J Virol. 2020. PMID: 32404529 Free PMC article.
-
Structural basis of receptor recognition by SARS-CoV-2.Nature. 2020 May;581(7807):221-224. doi: 10.1038/s41586-020-2179-y. Epub 2020 Mar 30. Nature. 2020. PMID: 32225175 Free PMC article.
-
Angiotensin-converting enzyme 2: The old door for new severe acute respiratory syndrome coronavirus 2 infection.Rev Med Virol. 2020 Sep;30(5):e2122. doi: 10.1002/rmv.2122. Epub 2020 Jun 30. Rev Med Virol. 2020. PMID: 32602627 Free PMC article. Review.
-
The SARS-CoV-2 Spike Glycoprotein as a Drug and Vaccine Target: Structural Insights into Its Complexes with ACE2 and Antibodies.Cells. 2020 Oct 22;9(11):2343. doi: 10.3390/cells9112343. Cells. 2020. PMID: 33105869 Free PMC article. Review.
Cited by
-
SARS-CoV 2 Infection (Covid-19) and Cardiovascular Disease in Africa: Health Care and Socio-Economic Implications.Glob Heart. 2021 Mar 15;16(1):18. doi: 10.5334/gh.829. Glob Heart. 2021. PMID: 33833942 Free PMC article. Review.
-
Vascular Endothelial Growth Factor Receptor 2: Molecular Mechanism and Therapeutic Potential in Preeclampsia Comorbidity with Human Immunodeficiency Virus and Severe Acute Respiratory Syndrome Coronavirus 2 Infections.Int J Mol Sci. 2022 Nov 9;23(22):13752. doi: 10.3390/ijms232213752. Int J Mol Sci. 2022. PMID: 36430232 Free PMC article. Review.
-
Serum β2-microglobulin levels in Coronavirus disease 2019 (Covid-19): Another prognosticator of disease severity?PLoS One. 2021 Mar 1;16(3):e0247758. doi: 10.1371/journal.pone.0247758. eCollection 2021. PLoS One. 2021. PMID: 33647017 Free PMC article.
-
The role of extracellular vesicles in COVID-19 virus infection.Infect Genet Evol. 2020 Nov;85:104422. doi: 10.1016/j.meegid.2020.104422. Epub 2020 Jun 13. Infect Genet Evol. 2020. PMID: 32544615 Free PMC article. Review.
-
Serum levels of ACE2 are higher in patients with obesity and diabetes.Obes Sci Pract. 2020 Dec 16;7(2):239-243. doi: 10.1002/osp4.472. eCollection 2021 Apr. Obes Sci Pract. 2020. PMID: 33841894 Free PMC article.
References
-
- Marra MA, Jones SJM, Astell CR, Holt RA, Brooks-Wilson A, Butterfield YSN, Khattra J, Asano JK, Barber SA, Chan SY, Cloutier A, Coughlin SM, Freeman D, Girn N, Griffith OL, Leach SR, Mayo M, McDonald H, Montgomery SB, Pandoh PK, Petrescu AS, Robertson AG, Schein JE, Siddiqui A, Smailus DE, Stott JM, Yang GS, Plummer F, Andonov A, Artsob H, Bastien N, Bernard K, Booth TF, Bowness D, Czub M, Drebot M, Fernando L, Flick R, Garbutt M, Gray M, Grolla A, Jones S, Feldmann H, Meyers A, Kabani A, Li Y, Normand S, Stroher U, Tipples GA, Tyler S, Vogrig R, Ward D, Watson B, Brunham RC, Krajden M, Petric M, Skowronski DM, Upton C, Roper RL. 2003. The genome sequence of the SARS-associated coronavirus. Science 300:1399–1404. doi:10.1126/science.1085953. - DOI - PubMed
-
- Peiris JSM, SARS Study Group, Lai ST, Poon LLM, Guan Y, Yam LYC, Lim W, Nicholls J, Yee WKS, Yan WW, Cheung MT, Cheng VCC, Chan KH, Tsang DNC, Yung RWH, Ng TK, Yuen KY. 2003. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 361:1319–1325. doi:10.1016/S0140-6736(03)13077-2. - DOI - PMC - PubMed
-
- Guan Y, Zheng BJ, He YQ, Liu XL, Zhuang ZX, Cheung CL, Luo SW, Li PH, Zhang LJ, Guan YJ, Butt KM, Wong KL, Chan KW, Lim W, Shortridge KF, Yuen KY, Peiris JSM, Poon L. 2003. Isolation and characterization of viruses related to the SARS coronavirus from animals in Southern China. Science 302:276–278. doi:10.1126/science.1087139. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
