

Terminal neuron localization to the upper cortical plate is controlled by the transcription factor NEUROD2
- PMID: 31873146
- PMCID: PMC6927953
- DOI: 10.1038/s41598-019-56171-x
Terminal neuron localization to the upper cortical plate is controlled by the transcription factor NEUROD2
Abstract
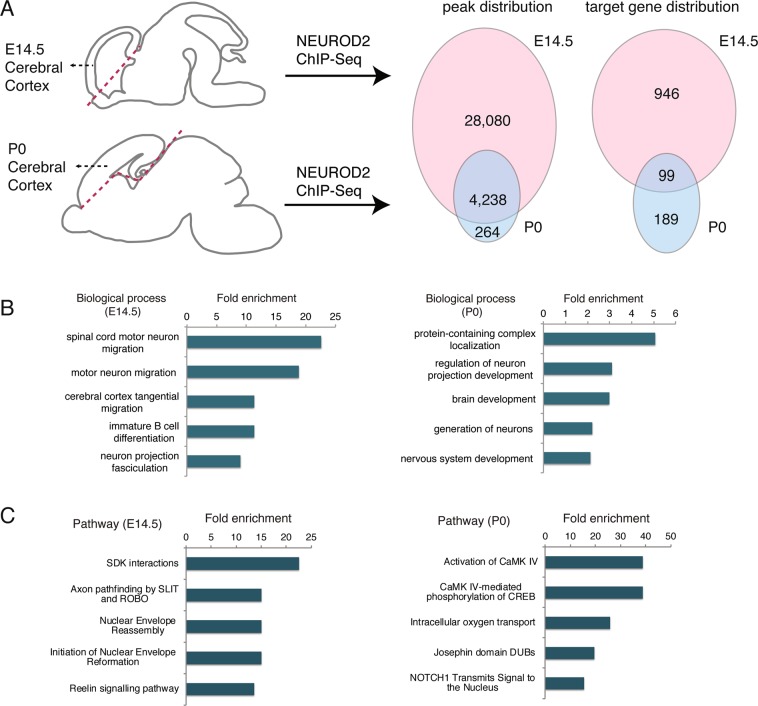
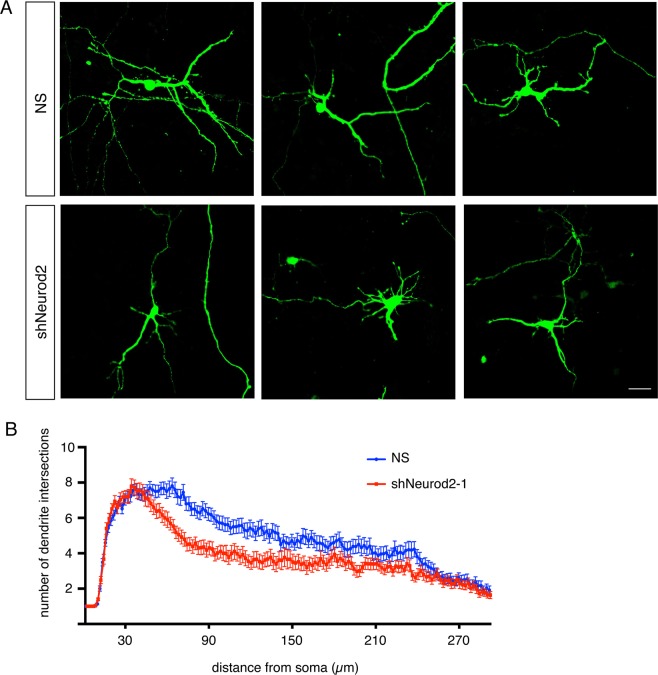
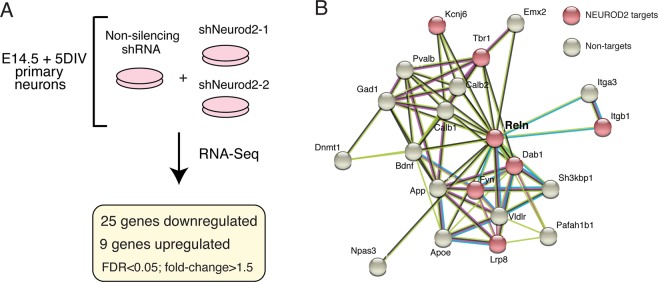
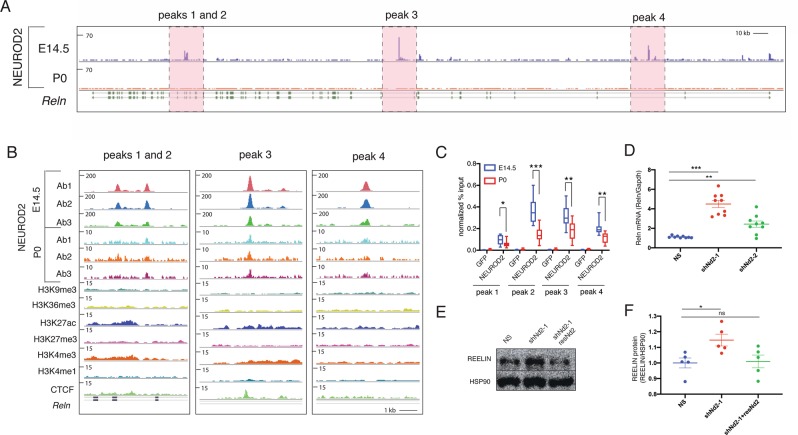
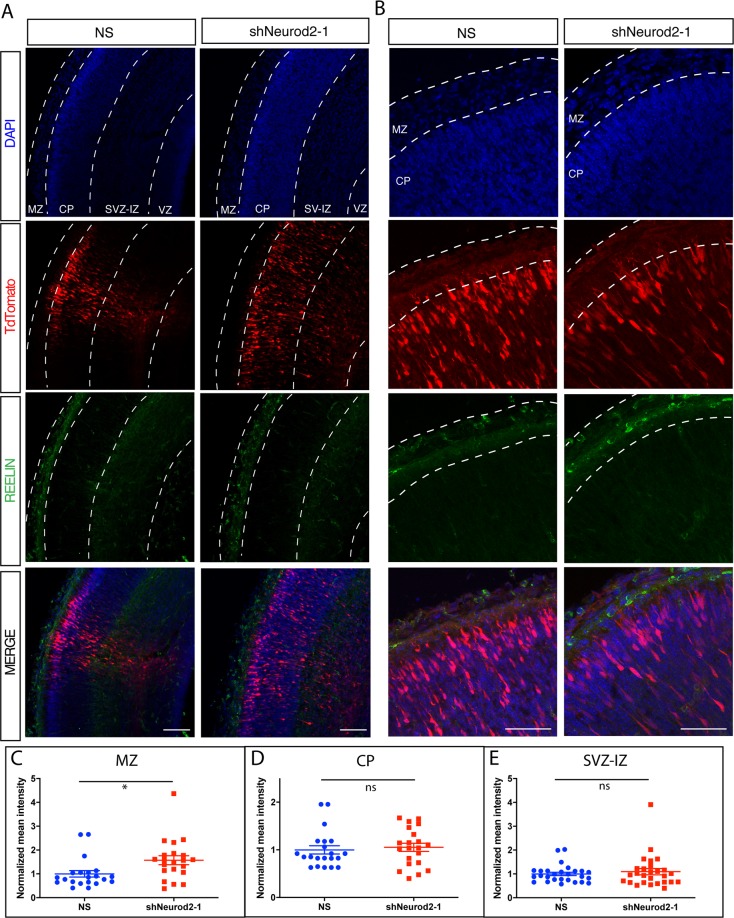
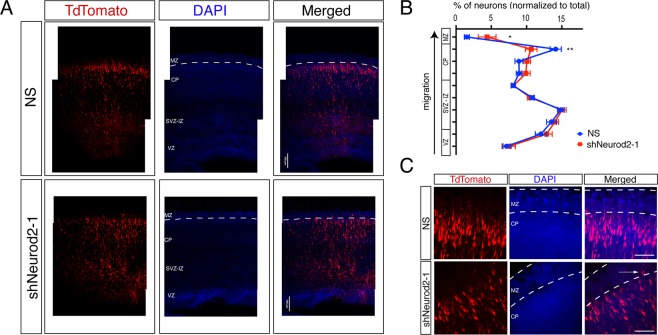
Excitatory neurons of the mammalian cerebral cortex are organized into six functional layers characterized by unique patterns of connectivity, as well as distinctive physiological and morphological properties. Cortical layers appear after a highly regulated migration process in which cells move from the deeper, proliferative zone toward the superficial layers. Importantly, defects in this radial migration process have been implicated in neurodevelopmental and psychiatric diseases. Here we report that during the final stages of migration, transcription factor Neurogenic Differentiation 2 (Neurod2) contributes to terminal cellular localization within the cortical plate. In mice, in utero knockdown of Neurod2 resulted in reduced numbers of neurons localized to the uppermost region of the developing cortex, also termed the primitive cortical zone. Our ChIP-Seq and RNA-Seq analyses of genes regulated by NEUROD2 in the developing cortex identified a number of key target genes with known roles in Reelin signaling, a critical regulator of neuronal migration. Our focused analysis of regulation of the Reln gene, encoding the extracellular ligand REELIN, uncovered NEUROD2 binding to conserved E-box elements in multiple introns. Furthermore, we demonstrate that knockdown of NEUROD2 in primary cortical neurons resulted in a strong increase in Reln gene expression at the mRNA level, as well as a slight upregulation at the protein level. These data reveal a new role for NEUROD2 during the late stages of neuronal migration, and our analysis of its genomic targets offers new genes with potential roles in cortical lamination.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Genome-wide target analysis of NEUROD2 provides new insights into regulation of cortical projection neuron migration and differentiation.BMC Genomics. 2015 Sep 5;16:681. doi: 10.1186/s12864-015-1882-9. BMC Genomics. 2015. PMID: 26341353 Free PMC article.
-
Analysis of Reelin signaling and neurodevelopmental trajectory in primary cultured cortical neurons with RELN deletion identified in schizophrenia.Neurochem Int. 2021 Mar;144:104954. doi: 10.1016/j.neuint.2020.104954. Epub 2021 Jan 1. Neurochem Int. 2021. PMID: 33388358
-
Synaptic activity suppresses expression of neurogenic differentiation factor 2 in an NMDA receptor-dependent manner.Synapse. 2017 Sep;71(9). doi: 10.1002/syn.21986. Epub 2017 Jun 2. Synapse. 2017. PMID: 28524267
-
Molecular pathways controlling the sequential steps of cortical projection neuron migration.Adv Exp Med Biol. 2014;800:1-24. doi: 10.1007/978-94-007-7687-6_1. Adv Exp Med Biol. 2014. PMID: 24243097 Review.
-
Role of Cajal-Retzius and subplate neurons in cerebral cortical development.Semin Pediatr Neurol. 2002 Dec;9(4):302-8. doi: 10.1053/spen.2002.32506. Semin Pediatr Neurol. 2002. PMID: 12523554 Review.
Cited by
-
Disruption of NEUROD2 causes a neurodevelopmental syndrome with autistic features via cell-autonomous defects in forebrain glutamatergic neurons.Mol Psychiatry. 2021 Nov;26(11):6125-6148. doi: 10.1038/s41380-021-01179-x. Epub 2021 Jun 29. Mol Psychiatry. 2021. PMID: 34188164 Free PMC article.
-
The Prevailing Role of Topoisomerase 2 Beta and its Associated Genes in Neurons.Mol Neurobiol. 2021 Dec;58(12):6443-6459. doi: 10.1007/s12035-021-02561-0. Epub 2021 Sep 21. Mol Neurobiol. 2021. PMID: 34546528 Review.
-
Neuro2D Lies at the Nexus of Autism, Epilepsy, and Intellectual Disabilities.Epilepsy Curr. 2022 Apr 4;22(2):132-134. doi: 10.1177/15357597211073424. eCollection 2022 Mar-Apr. Epilepsy Curr. 2022. PMID: 35444512 Free PMC article. No abstract available.
-
New Insights Into the Intricacies of Proneural Gene Regulation in the Embryonic and Adult Cerebral Cortex.Front Mol Neurosci. 2021 Feb 15;14:642016. doi: 10.3389/fnmol.2021.642016. eCollection 2021. Front Mol Neurosci. 2021. PMID: 33658912 Free PMC article. Review.
-
Massively parallel reporter perturbation assays uncover temporal regulatory architecture during neural differentiation.Nat Commun. 2022 Mar 21;13(1):1504. doi: 10.1038/s41467-022-28659-0. Nat Commun. 2022. PMID: 35315433 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
