

Early antiretroviral therapy in neonates with HIV-1 infection restricts viral reservoir size and induces a distinct innate immune profile
- PMID: 31776292
- PMCID: PMC8397898
- DOI: 10.1126/scitranslmed.aax7350
Early antiretroviral therapy in neonates with HIV-1 infection restricts viral reservoir size and induces a distinct innate immune profile
Abstract
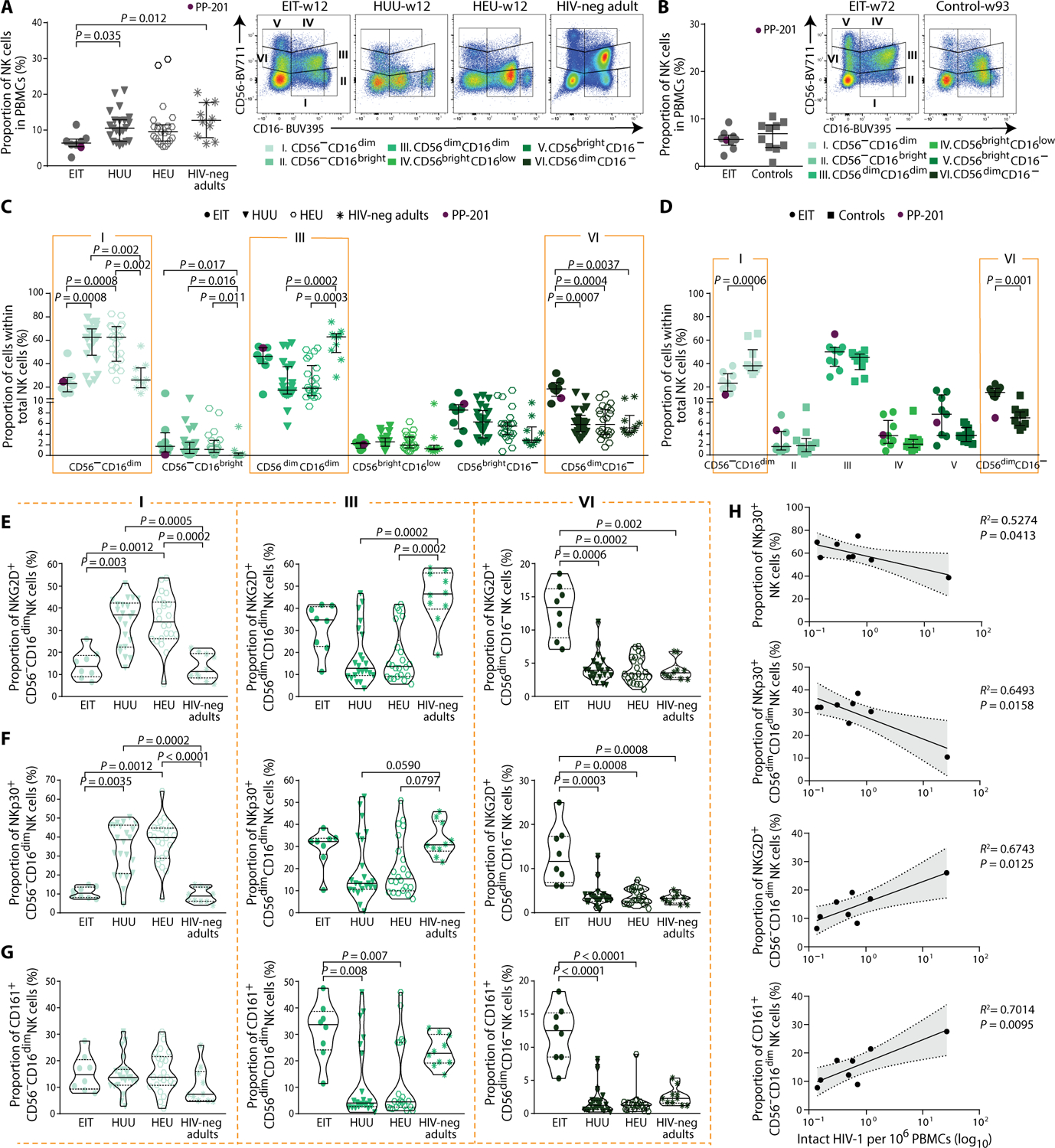
Neonatal HIV-1 infection is associated with rapidly progressive and frequently fatal immune deficiency if left untreated. Immediate institution of antiretroviral therapy (ART), ideally within hours after birth, may restrict irreversible damage to the developing neonatal immune system and possibly provide opportunities for facilitating drug-free viral control during subsequent treatment interruptions. However, the virological and immunological effects of ART initiation within hours after delivery have not been systematically investigated. We examined a unique cohort of neonates with HIV-1 infection from Botswana who started ART shortly after birth and were followed longitudinally for about 2 years in comparison to control infants started on treatment during the first year after birth. We demonstrate multiple clear benefits of rapid antiretroviral initiation, including an extremely small reservoir of intact proviral sequences, a reduction in abnormal T cell immune activation, a more polyfunctional HIV-1-specific T cell response, and an innate immune profile that displays distinct features of improved antiviral activity and is associated with intact proviral reservoir size. Together, these data offer rare insight into the evolutionary dynamics of viral reservoir establishment in neonates and provide strong empirical evidence supporting the immediate initiation of ART for neonates with HIV-1 infection.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures




Similar articles
-
Immune correlates of HIV-1 reservoir cell decline in early-treated infants.Cell Rep. 2022 Jul 19;40(3):111126. doi: 10.1016/j.celrep.2022.111126. Cell Rep. 2022. PMID: 35858580 Free PMC article.
-
Establishment and Replenishment of the Viral Reservoir in Perinatally HIV-1-infected Children Initiating Very Early Antiretroviral Therapy.Clin Infect Dis. 2015 Oct 1;61(7):1169-78. doi: 10.1093/cid/civ456. Epub 2015 Jun 10. Clin Infect Dis. 2015. PMID: 26063721 Free PMC article.
-
Genetic Diversity, Compartmentalization, and Age of HIV Proviruses Persisting in CD4+ T Cell Subsets during Long-Term Combination Antiretroviral Therapy.J Virol. 2020 Feb 14;94(5):e01786-19. doi: 10.1128/JVI.01786-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31776273 Free PMC article.
-
HIV-1 Persistence in Children during Suppressive ART.Viruses. 2021 Jun 12;13(6):1134. doi: 10.3390/v13061134. Viruses. 2021. PMID: 34204740 Free PMC article. Review.
-
Residual Proviral Reservoirs: A High Risk for HIV Persistence and Driving Forces for Viral Rebound after Analytical Treatment Interruption.Viruses. 2021 Feb 21;13(2):335. doi: 10.3390/v13020335. Viruses. 2021. PMID: 33670027 Free PMC article. Review.
Cited by
-
Differences in inducibility of the latent HIV reservoir in perinatal and adult infection.JCI Insight. 2020 Feb 27;5(4):e134105. doi: 10.1172/jci.insight.134105. JCI Insight. 2020. PMID: 31999647 Free PMC article.
-
Interventions during Early Infection: Opening a Window for an HIV Cure?Viruses. 2024 Oct 9;16(10):1588. doi: 10.3390/v16101588. Viruses. 2024. PMID: 39459922 Free PMC article. Review.
-
T cell immune discriminants of HIV reservoir size in a pediatric cohort of perinatally infected individuals.PLoS Pathog. 2021 Apr 26;17(4):e1009533. doi: 10.1371/journal.ppat.1009533. eCollection 2021 Apr. PLoS Pathog. 2021. PMID: 33901266 Free PMC article.
-
HIV-1 reservoir size after neonatal antiretroviral therapy and the potential to evaluate antiretroviral-therapy-free remission (IMPAACT P1115): a phase 1/2 proof-of-concept study.Lancet HIV. 2024 Jan;11(1):e20-e30. doi: 10.1016/S2352-3018(23)00236-9. Epub 2023 Dec 4. Lancet HIV. 2024. PMID: 38061376 Free PMC article. Clinical Trial.
-
Initiation of Antiretroviral Therapy during Primary HIV Infection: Effects on the Latent HIV Reservoir, Including on Analytic Treatment Interruptions.AIDS Rev. 2020 Oct 26;23(1):28-39. doi: 10.24875/AIDSRev.20000001. AIDS Rev. 2020. PMID: 33105471 Free PMC article.
References
-
- Siegfried N, van der Merwe L, Brocklehurst P, Sint TT, Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection. Cochrane Database Syst. Rev 2011, CD003510 (2011). - PubMed
-
- Newell ML, Coovadia H, Cortina-Borja M, Rollins N, Gaillard P; Francois Dabis for the Ghent International AIDS Society (IAS) working group on HIV infection in women and children, Mortality of infected and uninfected infants born to HIV-infected mothers in Africa: A pooled analysis. Lancet 364, 1236–1243 (2004). - PubMed
-
- World Health Organization, Antiretroviral Therapy for HIV Infection in Infants and Children: Towards Universal Access—Recommendations for a Public Health Approach: 2010 Revision (World Health Organization, 2010); www.who.int/hiv/pub/paediatric/infants2010/en/index.html. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 AI135940/AI/NIAID NIH HHS/United States
- R01 HL134539/HL/NHLBI NIH HHS/United States
- U01 AI114235/AI/NIAID NIH HHS/United States
- 107752/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- R33 DA047034/DA/NIDA NIH HHS/United States
- R61 DA047034/DA/NIDA NIH HHS/United States
- R01 AI130005/AI/NIAID NIH HHS/United States
- R33 AI116228/AI/NIAID NIH HHS/United States
- R33 AI122377/AI/NIAID NIH HHS/United States
- R21 AI124776/AI/NIAID NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 AI120008/AI/NIAID NIH HHS/United States
- R21 AI116228/AI/NIAID NIH HHS/United States
- R21 AI120850/AI/NIAID NIH HHS/United States
- R21 AI122377/AI/NIAID NIH HHS/United States
- R01 AI078799/AI/NIAID NIH HHS/United States
- U01 AI117841/AI/NIAID NIH HHS/United States
- R01 DK120387/DK/NIDDK NIH HHS/United States
- R56 AI125109/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
