

Evolution of placental invasion and cancer metastasis are causally linked
- PMID: 31768023
- PMCID: PMC7340496
- DOI: 10.1038/s41559-019-1046-4
Evolution of placental invasion and cancer metastasis are causally linked
Erratum in
-
Author Correction: Evolution of placental invasion and cancer metastasis are causally linked.Nat Ecol Evol. 2020 Feb;4(2):280. doi: 10.1038/s41559-020-1100-2. Nat Ecol Evol. 2020. PMID: 31949306
Abstract
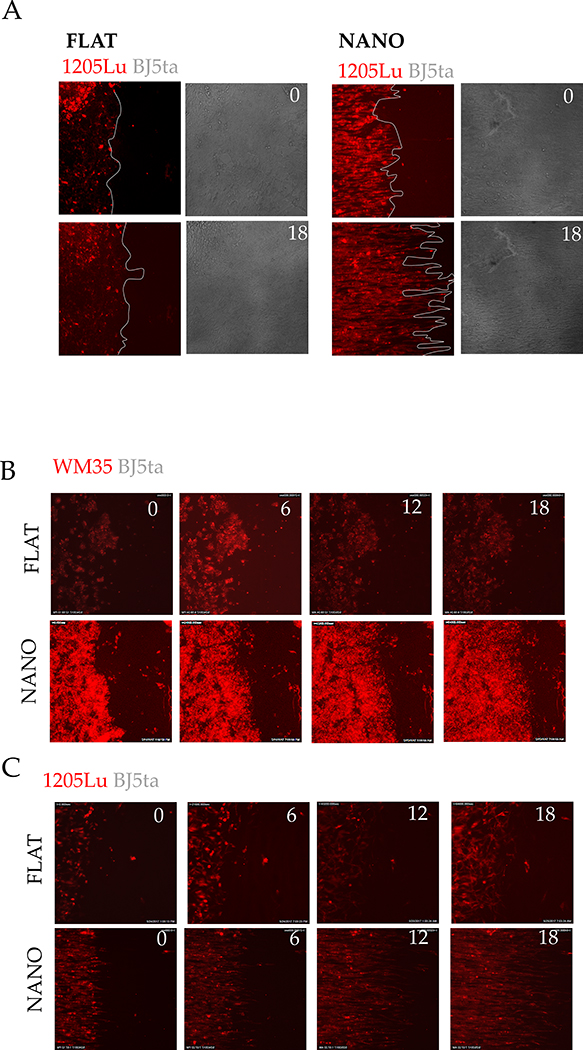
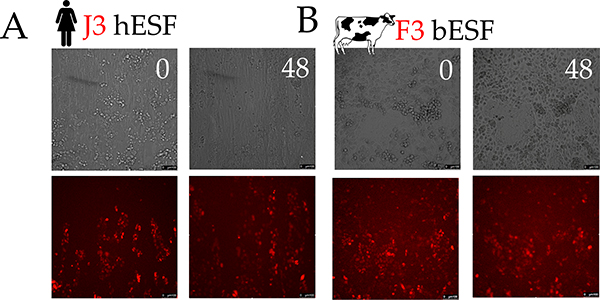
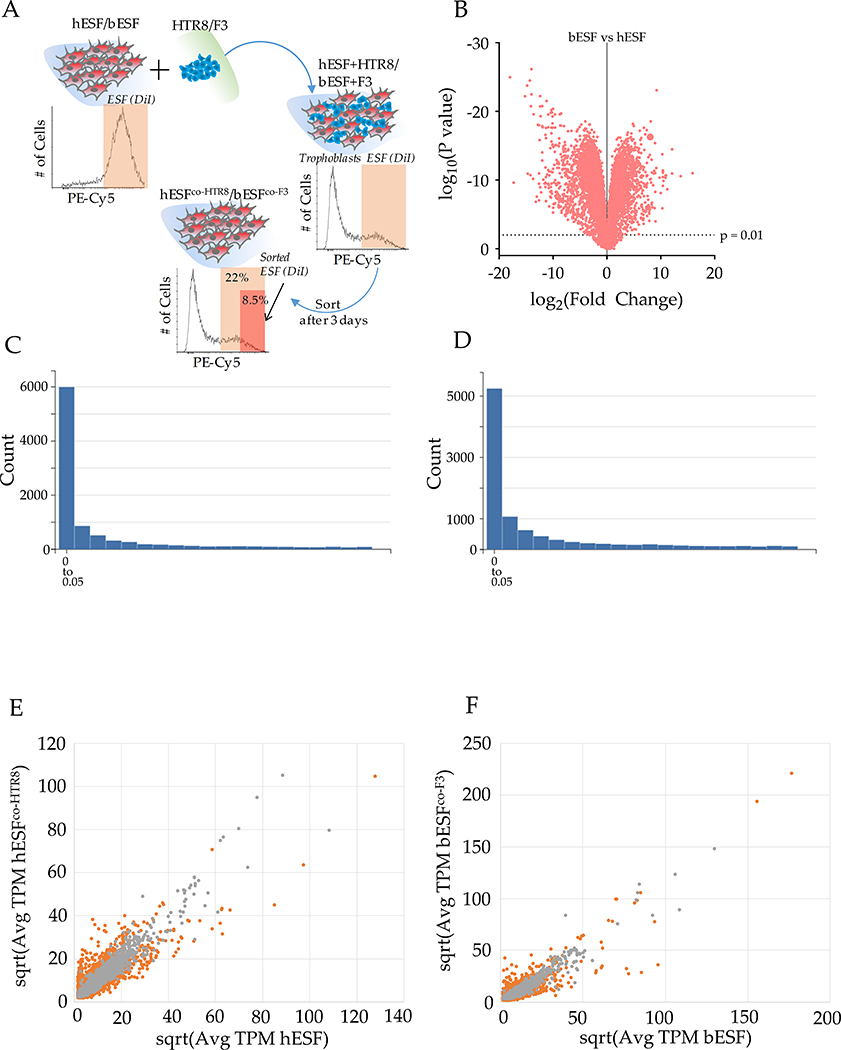
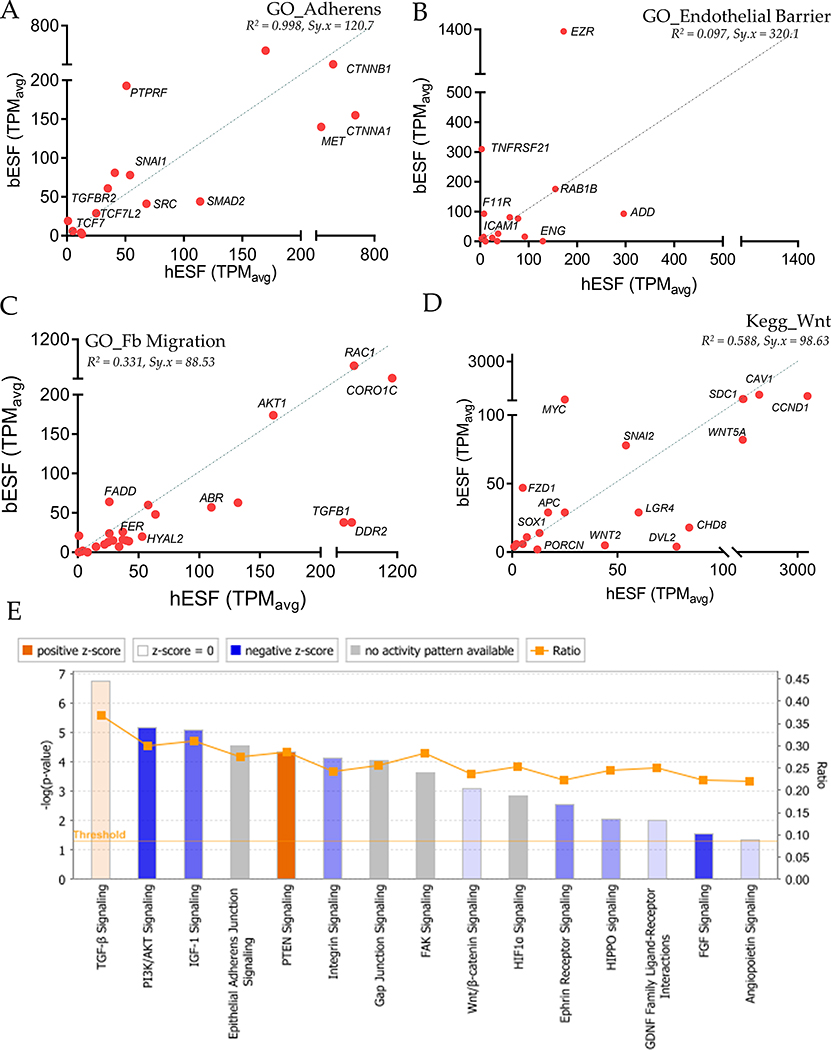
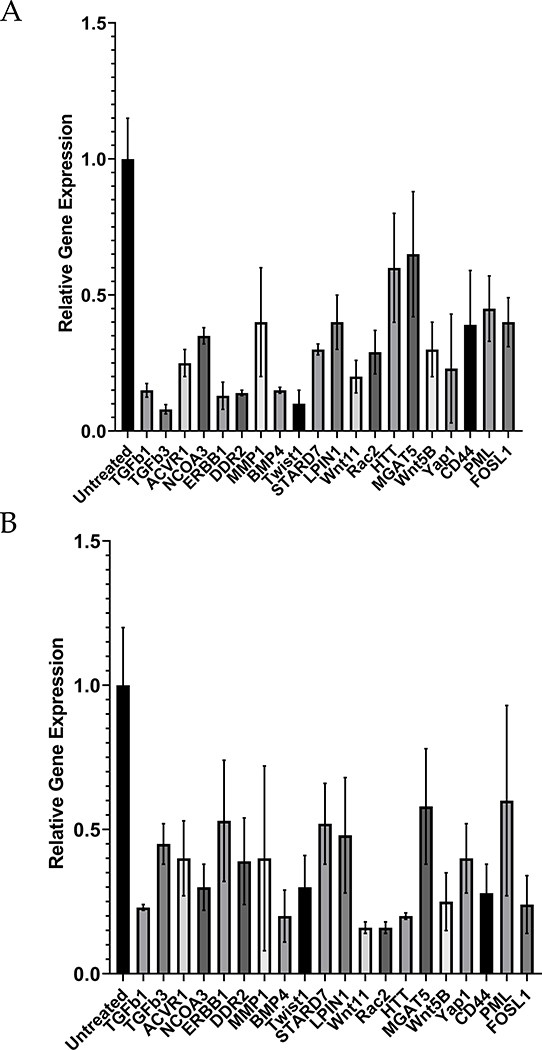
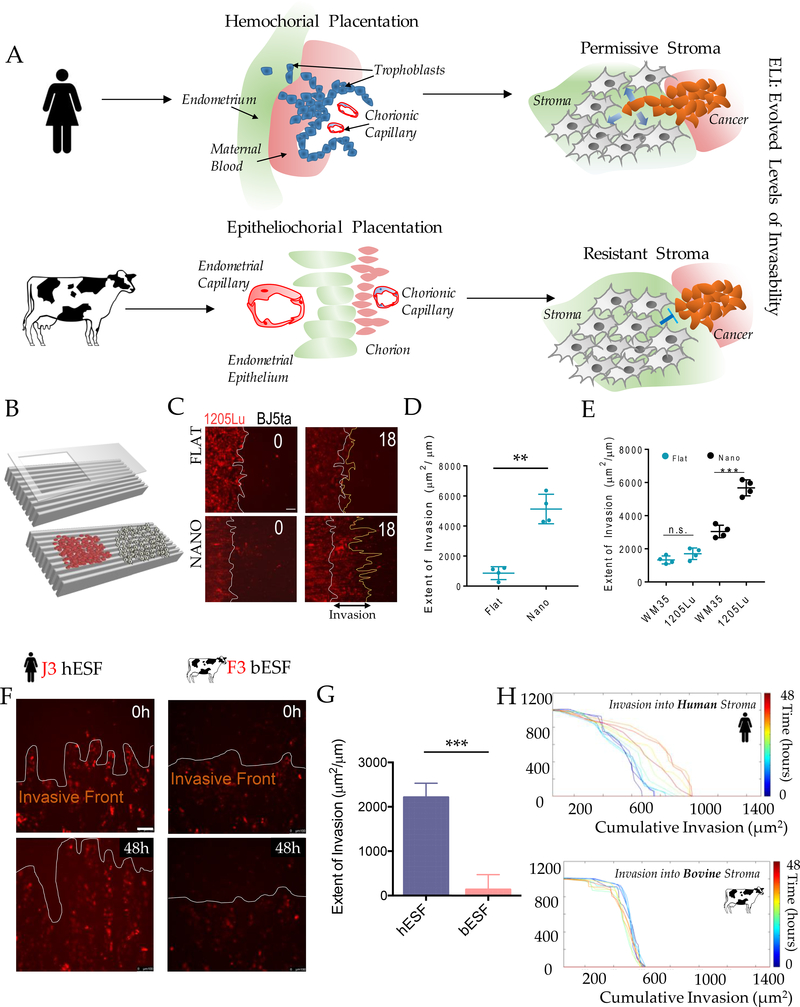
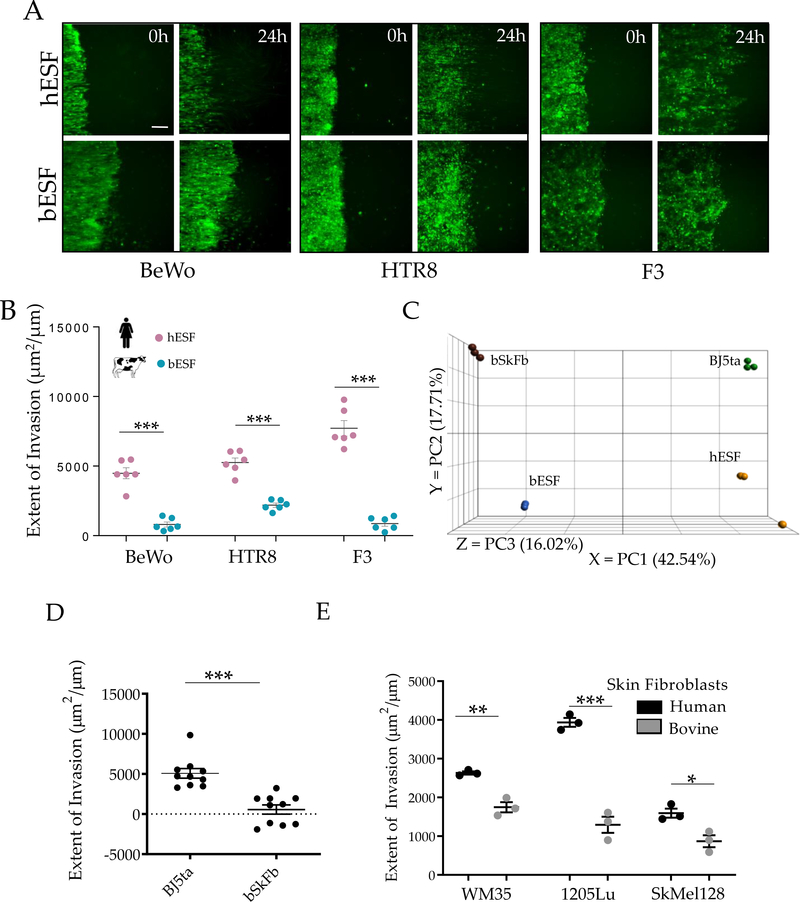
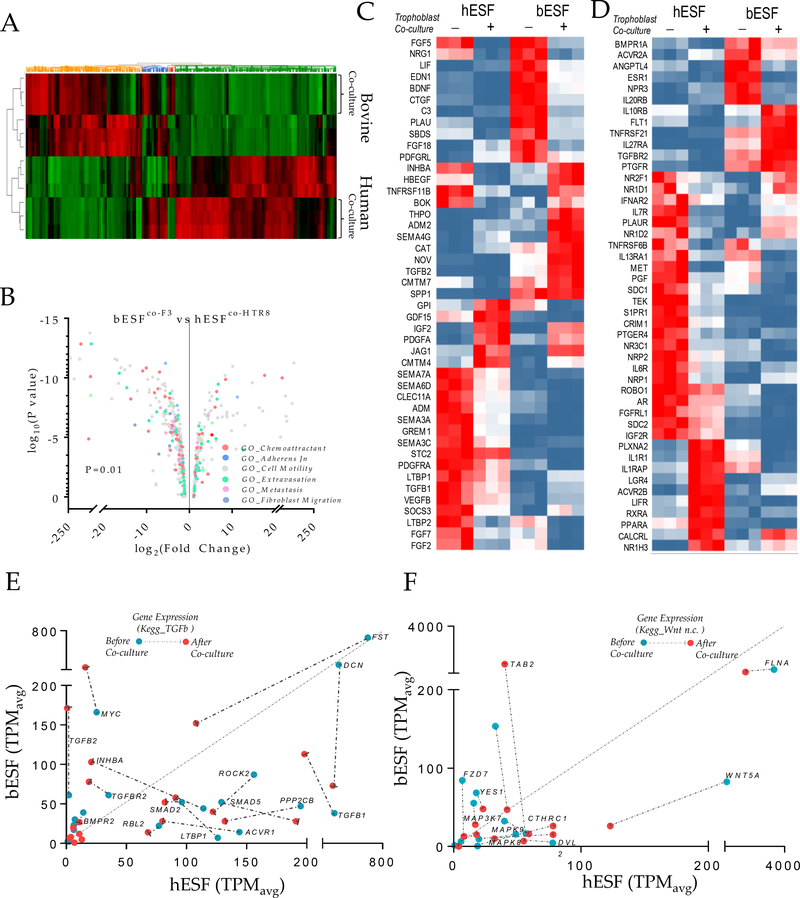
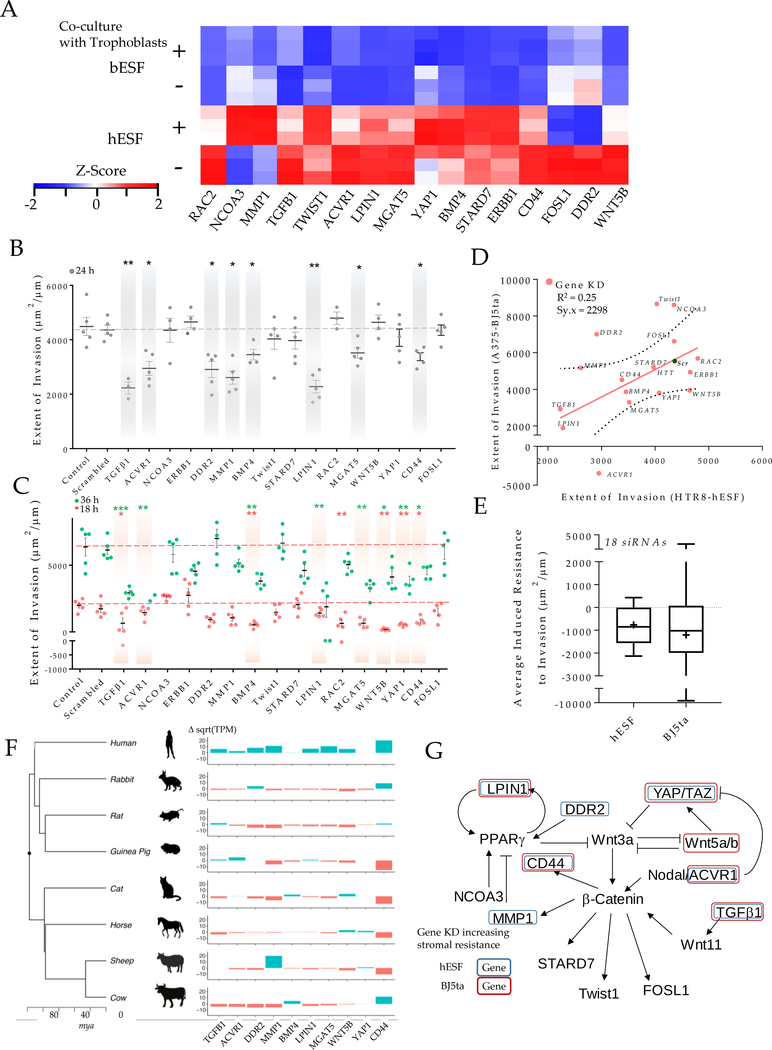
Among mammals, placental invasion is correlated with vulnerability to malignancy. Animals with more invasive placentation (for example, humans) are more vulnerable to malignancy. To explain this correlation, we propose the hypothesis of 'Evolved Levels of Invasibility' proposing that the evolution of invasibility of stromal tissue affects both placental and cancer invasion. We provide evidence for this using an in vitro model. We find that bovine endometrial and skin fibroblasts are more resistant to invasion than are their human counterparts. Gene expression profiling identified genes with high expression in human but not in bovine fibroblasts. Knocking down a subset of them in human fibroblasts leads to stronger resistance to cancer cell invasion. Identifying the evolutionary determinants of stromal invasibility can provide important insights to develop rational antimetastatic therapeutics.
Conflict of interest statement
Figures
Similar articles
-
Evolved Resistance to Placental Invasion Secondarily Confers Increased Survival in Melanoma Patients.J Clin Med. 2021 Feb 5;10(4):595. doi: 10.3390/jcm10040595. J Clin Med. 2021. PMID: 33562461 Free PMC article.
-
Tracing the cis-regulatory changes underlying the endometrial control of placental invasion.Proc Natl Acad Sci U S A. 2022 Feb 8;119(6):e2111256119. doi: 10.1073/pnas.2111256119. Proc Natl Acad Sci U S A. 2022. PMID: 35110402 Free PMC article.
-
Genetic recapitulation of human pre-eclampsia risk during convergent evolution of reduced placental invasiveness in eutherian mammals.Philos Trans R Soc Lond B Biol Sci. 2015 Mar 5;370(1663):20140069. doi: 10.1098/rstb.2014.0069. Philos Trans R Soc Lond B Biol Sci. 2015. PMID: 25602073 Free PMC article.
-
Malignant cancer and invasive placentation: A case for positive pleiotropy between endometrial and malignancy phenotypes.Evol Med Public Health. 2014 Oct 15;2014(1):136-45. doi: 10.1093/emph/eou022. Evol Med Public Health. 2014. PMID: 25324490 Free PMC article. Review.
-
What was the ancestral function of decidual stromal cells? A model for the evolution of eutherian pregnancy.Placenta. 2016 Apr;40:40-51. doi: 10.1016/j.placenta.2016.02.012. Epub 2016 Feb 23. Placenta. 2016. PMID: 27016782 Review.
Cited by
-
Evolution of higher mesenchymal CD44 expression in the human lineage: A gene linked to cancer malignancy.Evol Med Public Health. 2022 Aug 30;10(1):447-462. doi: 10.1093/emph/eoac036. eCollection 2022. Evol Med Public Health. 2022. PMID: 36148042 Free PMC article.
-
BAP1/ASXL complex modulation regulates epithelial-mesenchymal transition during trophoblast differentiation and invasion.Elife. 2021 Jun 25;10:e63254. doi: 10.7554/eLife.63254. Elife. 2021. PMID: 34170818 Free PMC article.
-
A common allele increases endometrial Wnt4 expression, with antagonistic implications for pregnancy, reproductive cancers, and endometriosis.Nat Commun. 2024 Feb 12;15(1):1152. doi: 10.1038/s41467-024-45338-4. Nat Commun. 2024. PMID: 38346980 Free PMC article.
-
Gene expression phylogenies and ancestral transcriptome reconstruction resolves major transitions in the origins of pregnancy.Elife. 2022 Jun 30;11:e74297. doi: 10.7554/eLife.74297. Elife. 2022. PMID: 35770963 Free PMC article.
-
Evolved Resistance to Placental Invasion Secondarily Confers Increased Survival in Melanoma Patients.J Clin Med. 2021 Feb 5;10(4):595. doi: 10.3390/jcm10040595. J Clin Med. 2021. PMID: 33562461 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
