

Sumoylation of the DNA polymerase ε by the Smc5/6 complex contributes to DNA replication
- PMID: 31765372
- PMCID: PMC6876774
- DOI: 10.1371/journal.pgen.1008426
Sumoylation of the DNA polymerase ε by the Smc5/6 complex contributes to DNA replication
Abstract
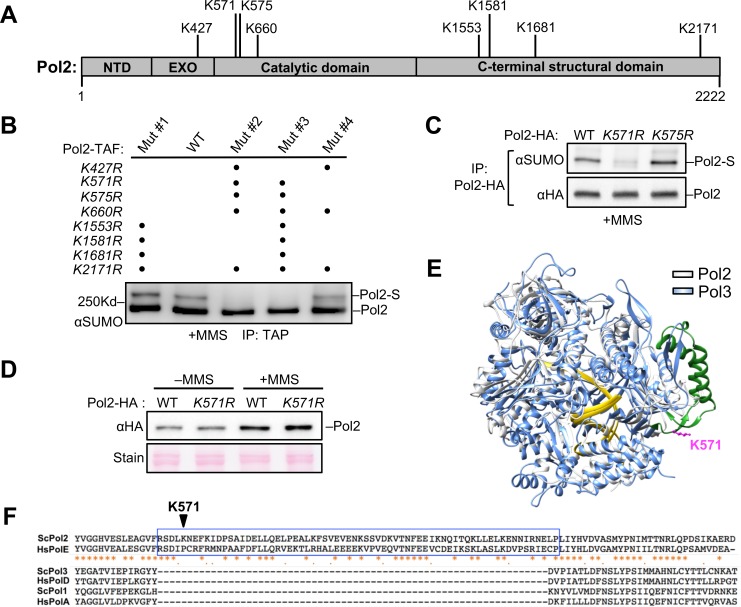
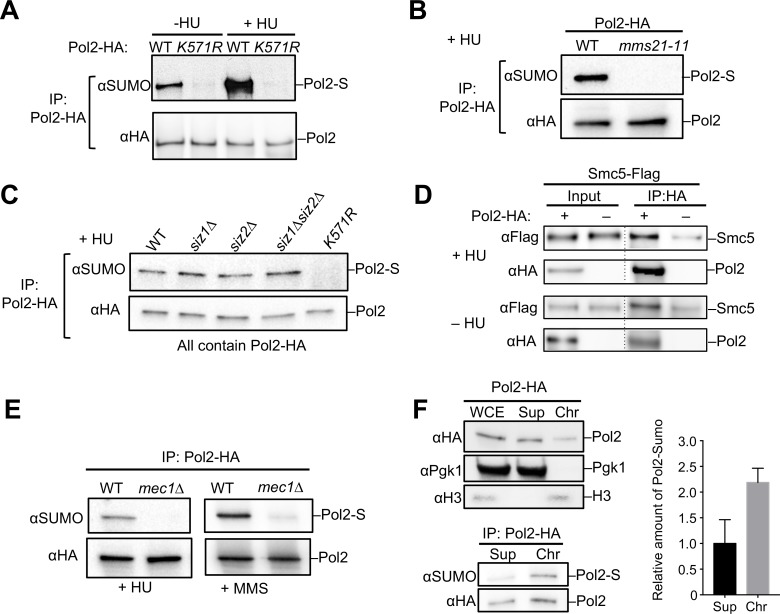
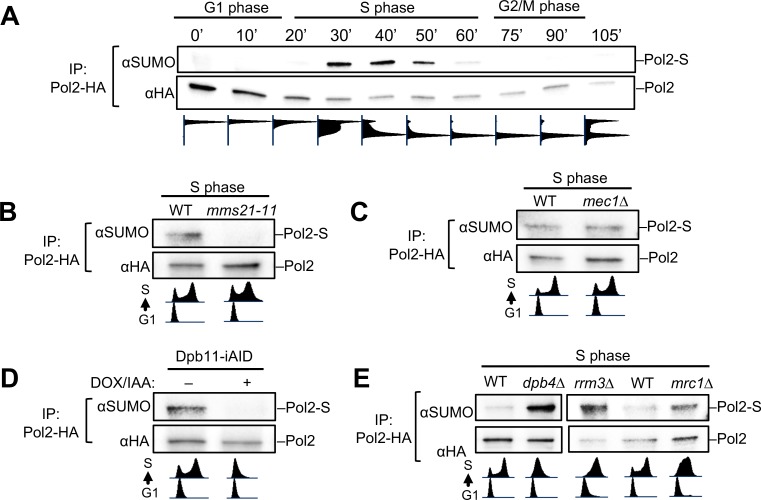
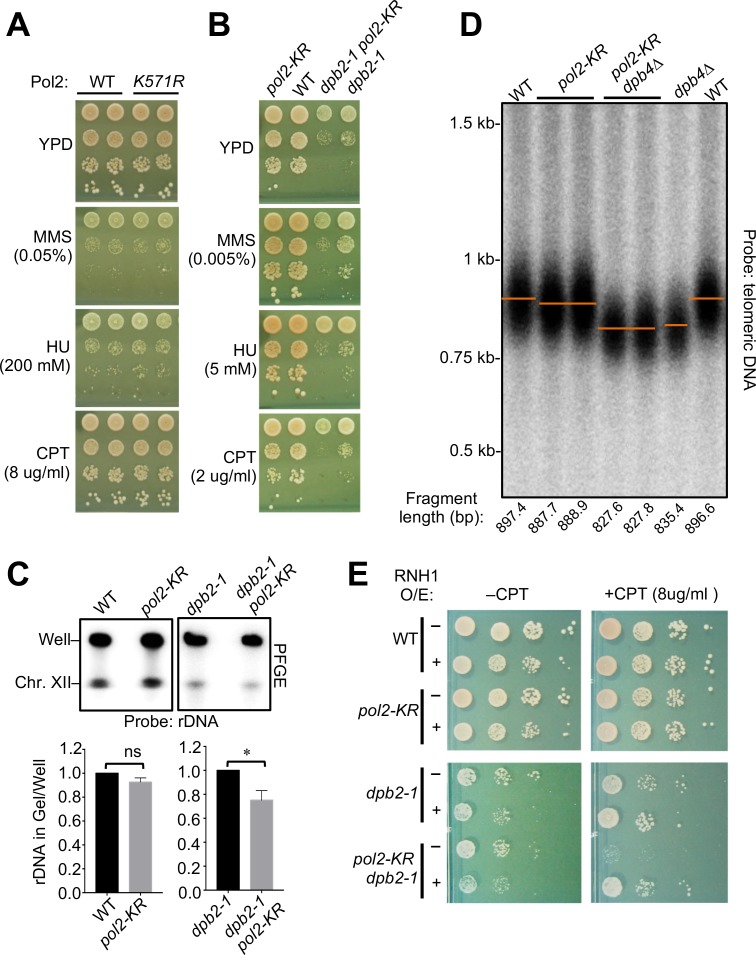
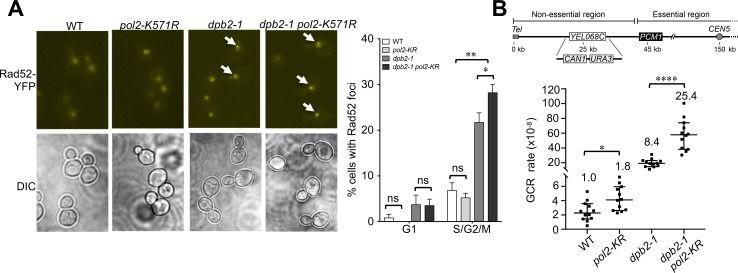
DNA polymerase epsilon (Pol ε) is critical for genome duplication, but little is known about how post-translational modification regulates its function. Here we report that the Pol ε catalytic subunit Pol2 in yeast is sumoylated at a single lysine within a catalytic domain insertion uniquely possessed by Pol2 family members. We found that Pol2 sumoylation occurs specifically in S phase and is increased under conditions of replication fork blockade. Analyses of the genetic requirements of this modification indicate that Pol2 sumoylation is associated with replication fork progression and dependent on the Smc5/6 SUMO ligase known to promote DNA synthesis. Consistently, the pol2 sumoylation mutant phenotype suggests impaired replication progression and increased levels of gross chromosomal rearrangements. Our findings thus indicate a direct role for SUMO in Pol2-mediated DNA synthesis and a molecular basis for Smc5/6-mediated regulation of genome stability.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The S phase checkpoint promotes the Smc5/6 complex dependent SUMOylation of Pol2, the catalytic subunit of DNA polymerase ε.PLoS Genet. 2019 Nov 25;15(11):e1008427. doi: 10.1371/journal.pgen.1008427. eCollection 2019 Nov. PLoS Genet. 2019. PMID: 31765407 Free PMC article.
-
Sumoylation of Smc5 Promotes Error-free Bypass at Damaged Replication Forks.Cell Rep. 2019 Dec 3;29(10):3160-3172.e4. doi: 10.1016/j.celrep.2019.10.123. Cell Rep. 2019. PMID: 31801080
-
Genetic evidence for functional interaction of Smc5/6 complex and Top1 with spatial frequency of replication origins required for maintenance of chromosome stability.Curr Genet. 2017 Aug;63(4):765-776. doi: 10.1007/s00294-017-0680-0. Epub 2017 Feb 16. Curr Genet. 2017. PMID: 28204881
-
Smc5/6 complex regulates Sgs1 recombination functions.Curr Genet. 2017 Jun;63(3):381-388. doi: 10.1007/s00294-016-0648-5. Epub 2016 Sep 23. Curr Genet. 2017. PMID: 27664093 Free PMC article. Review.
-
The evolving tale of Pol2 function.Genes Dev. 2023 Feb 1;37(3-4):72-73. doi: 10.1101/gad.350527.123. Epub 2023 Feb 22. Genes Dev. 2023. PMID: 36813532 Free PMC article. Review.
Cited by
-
SMC5/6 Promotes Replication Fork Stability via Negative Regulation of the COP9 Signalosome.Int J Mol Sci. 2024 Jan 12;25(2):952. doi: 10.3390/ijms25020952. Int J Mol Sci. 2024. PMID: 38256025 Free PMC article.
-
Coordinating DNA Replication and Mitosis through Ubiquitin/SUMO and CDK1.Int J Mol Sci. 2021 Aug 16;22(16):8796. doi: 10.3390/ijms22168796. Int J Mol Sci. 2021. PMID: 34445496 Free PMC article. Review.
-
Virus-host protein co-expression networks reveal temporal organization and strategies of viral infection.iScience. 2023 Nov 16;26(12):108475. doi: 10.1016/j.isci.2023.108475. eCollection 2023 Dec 15. iScience. 2023. PMID: 38077135 Free PMC article.
-
DNA polymerase ε relies on a unique domain for efficient replisome assembly and strand synthesis.Nat Commun. 2020 May 15;11(1):2437. doi: 10.1038/s41467-020-16095-x. Nat Commun. 2020. PMID: 32415104 Free PMC article.
-
The SMC5/6 complex: folding chromosomes back into shape when genomes take a break.Nucleic Acids Res. 2024 Mar 21;52(5):2112-2129. doi: 10.1093/nar/gkae103. Nucleic Acids Res. 2024. PMID: 38375830 Free PMC article. Review.
References
-
- Hamatake RK, Hasegawa H, Clark AB, Bebenek K, Kunkel TA, Sugino A. Purification and characterization of DNA polymerase II from the yeast Saccharomyces cerevisiae. Identification of the catalytic core and a possible holoenzyme form of the enzyme. J Biol Chem. 1990;265(7):4072–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
