

Disruption of palmitate-mediated localization; a shared pathway of force and anesthetic activation of TREK-1 channels
- PMID: 31672538
- PMCID: PMC6907892
- DOI: 10.1016/j.bbamem.2019.183091
Disruption of palmitate-mediated localization; a shared pathway of force and anesthetic activation of TREK-1 channels
Abstract
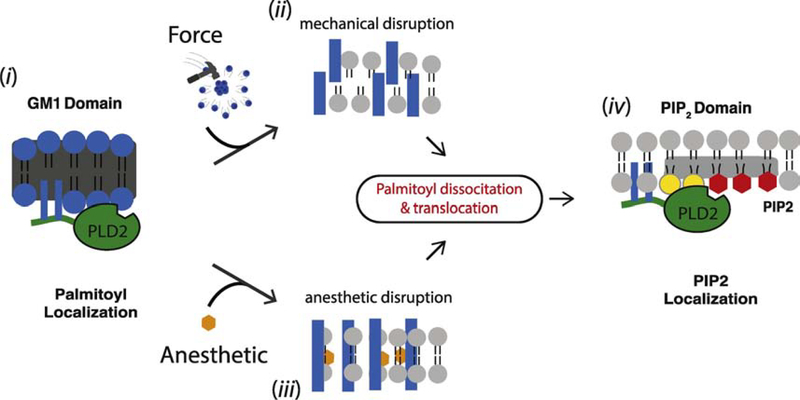
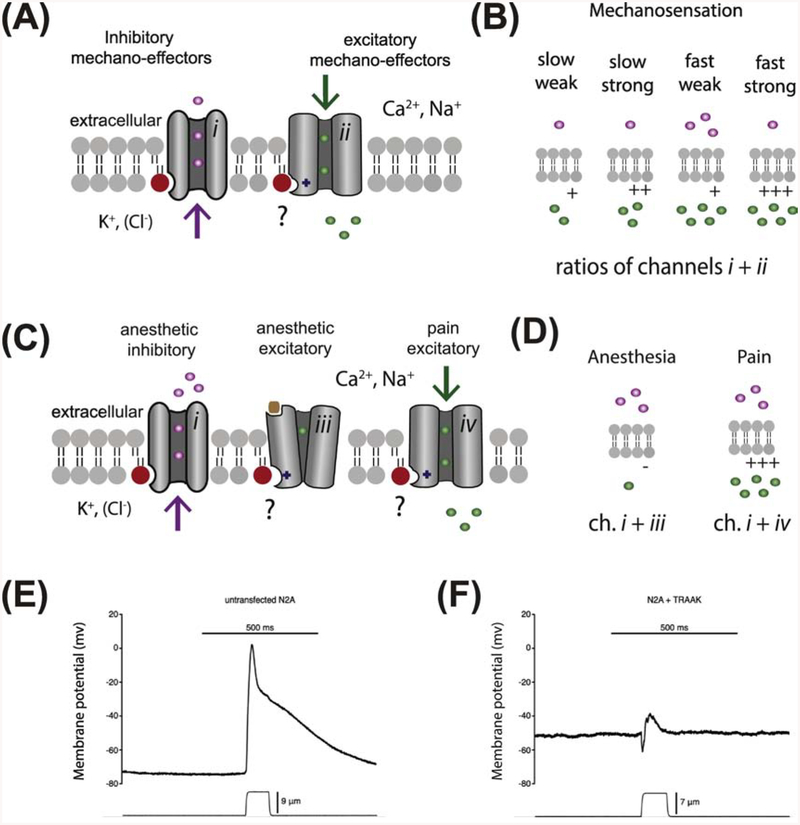
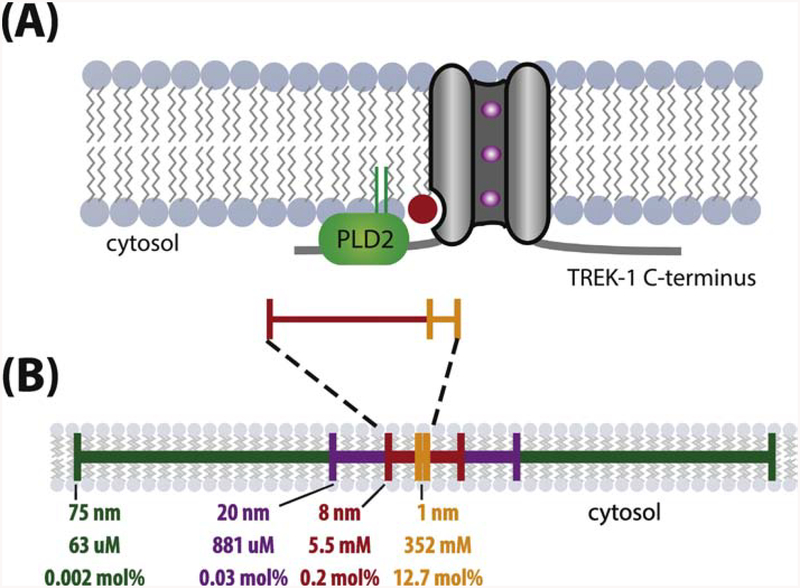
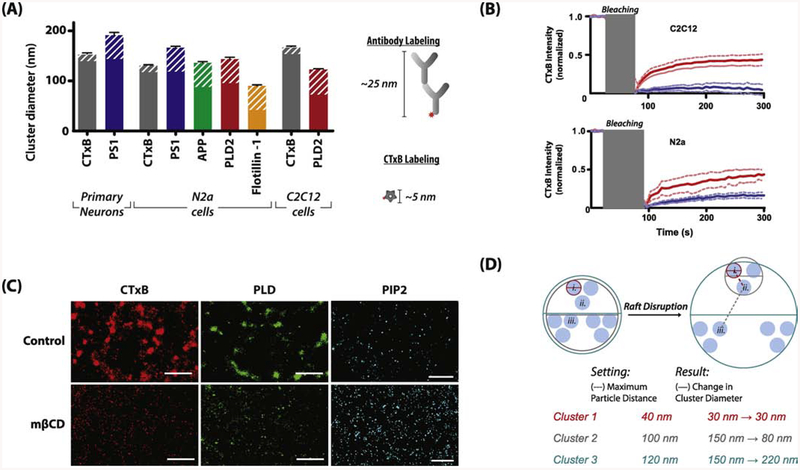
TWIK related K+ channel (TREK-1) is a mechano- and anesthetic sensitive channel that when activated attenuates pain and causes anesthesia. Recently the enzyme phospholipase D2 (PLD2) was shown to bind to the channel and generate a local high concentration of phosphatidic acid (PA), an anionic signaling lipid that gates TREK-1. In a biological membrane, the cell harnesses lipid heterogeneity (lipid compartments) to control gating of TREK-1 using palmitate-mediated localization of PLD2. Here we discuss the ability of mechanical force and anesthetics to disrupt palmitate-mediated localization of PLD2 giving rise to TREK-1's mechano- and anesthetic-sensitive properties. The likely consequences of this indirect lipid-based mechanism of activation are discussed in terms of a putative model for excitatory and inhibitory mechano-effectors and anesthetic sensitive ion channels in a biological context. Lastly, we discuss the ability of locally generated PA to reach mM concentrations near TREK-1 and the biophysics of localized signaling. Palmitate-mediated localization of PLD2 emerges as a central control mechanism of TREK-1 responding to mechanical force and anesthetic action. This article is part of a Special Issue entitled: Molecular biophysics of membranes and membrane proteins.
Keywords: Anesthesia; Lipid; Mechanosensation; Palmitoylation; Rafts; Super resolution microscopy.
Copyright © 2019 Elsevier B.V. All rights reserved.
Conflict of interest statement
Authors declare no conflicts of interests.
Figures
Similar articles
-
Polymodal Mechanism for TWIK-Related K+ Channel Inhibition by Local Anesthetic.Anesth Analg. 2019 Oct;129(4):973-982. doi: 10.1213/ANE.0000000000004216. Anesth Analg. 2019. PMID: 31124840
-
Studies on the mechanism of general anesthesia.Proc Natl Acad Sci U S A. 2020 Jun 16;117(24):13757-13766. doi: 10.1073/pnas.2004259117. Epub 2020 May 28. Proc Natl Acad Sci U S A. 2020. PMID: 32467161 Free PMC article.
-
Mechanical activation of TWIK-related potassium channel by nanoscopic movement and rapid second messenger signaling.Elife. 2024 Feb 26;12:RP89465. doi: 10.7554/eLife.89465. Elife. 2024. PMID: 38407149 Free PMC article.
-
Anesthetic Mechanisms: Synergistic Interactions With Lipid Rafts and Voltage-Gated Sodium Channels.Anesth Analg. 2024 Jul 1;139(1):92-106. doi: 10.1213/ANE.0000000000006738. Epub 2024 Jun 17. Anesth Analg. 2024. PMID: 37968836 Review.
-
Perspectives on the Two-Pore Domain Potassium Channel TREK-1 (TWIK-Related K(+) Channel 1). A Novel Therapeutic Target?J Med Chem. 2016 Jun 9;59(11):5149-57. doi: 10.1021/acs.jmedchem.5b00671. Epub 2015 Dec 14. J Med Chem. 2016. PMID: 26588045 Review.
Cited by
-
Roles of Membrane Domains in Integrin-Mediated Cell Adhesion.Int J Mol Sci. 2020 Aug 1;21(15):5531. doi: 10.3390/ijms21155531. Int J Mol Sci. 2020. PMID: 32752284 Free PMC article. Review.
-
Membrane phospholipids control gating of the mechanosensitive potassium leak channel TREK1.Nat Commun. 2023 Feb 25;14(1):1077. doi: 10.1038/s41467-023-36765-w. Nat Commun. 2023. PMID: 36841877 Free PMC article.
-
Negative Influence by the Force: Mechanically Induced Hyperpolarization via K2P Background Potassium Channels.Int J Mol Sci. 2021 Aug 23;22(16):9062. doi: 10.3390/ijms22169062. Int J Mol Sci. 2021. PMID: 34445768 Free PMC article. Review.
-
Hydroxychloroquine blocks SARS-CoV-2 entry into the endocytic pathway in mammalian cell culture.Commun Biol. 2022 Sep 14;5(1):958. doi: 10.1038/s42003-022-03841-8. Commun Biol. 2022. PMID: 36104427 Free PMC article.
-
GABA and astrocytic cholesterol determine the lipid environment of GABAAR in cultured cortical neurons.bioRxiv [Preprint]. 2024 Apr 29:2024.04.26.591395. doi: 10.1101/2024.04.26.591395. bioRxiv. 2024. PMID: 38746110 Free PMC article. Preprint.
References
-
- Honoré E, The neuronal background K2P channels: focus on TREK1, Nat. Rev. Neurosci, 8 (2007) 251–261. - PubMed
-
- Patel AJ, Honoré E, Lesage F, Fink M, Romey G, Lazdunski M, Inhalational anesthetics activate two-pore-domain background K+ channels, Nat. Neurosci, 2 (1999) 422–6. - PubMed
-
- Gruss M, Bushell TJ, Bright DP, Lieb WR, Mathie A, Franks NP, Two-pore-domain K+ channels are a novel target for the anesthetic gases xenon, nitrous oxide, and cyclopropane, Mol Pharmacol, 65 (2004) 443–452. - PubMed
-
- Dong YY, Pike ACW, Mackenzie A, Mcclenaghan C, Aryal P, Dong L, Quigley A, Grieben M, Goubin S, Mukhopadhyay S, Ruda GF, V Clausen M, Cao L, Brennan PE, a Burgess-brown N, Sansom MSP, Tucker SJ, Carpenter EP, K2P channel gating mechanisms revealed by structures of TREK-2 and a complex with Prozac, Science, 347 (2015) 1256–9. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
