

RNA-dependent Amplification of Mammalian mRNA Encoding Extracellullar Matrix Proteins: Identification of Chimeric RNA Intermediates for α1, β1, and γ1 Chains of Laminin
- PMID: 31663081
- PMCID: PMC6818727
RNA-dependent Amplification of Mammalian mRNA Encoding Extracellullar Matrix Proteins: Identification of Chimeric RNA Intermediates for α1, β1, and γ1 Chains of Laminin
Abstract
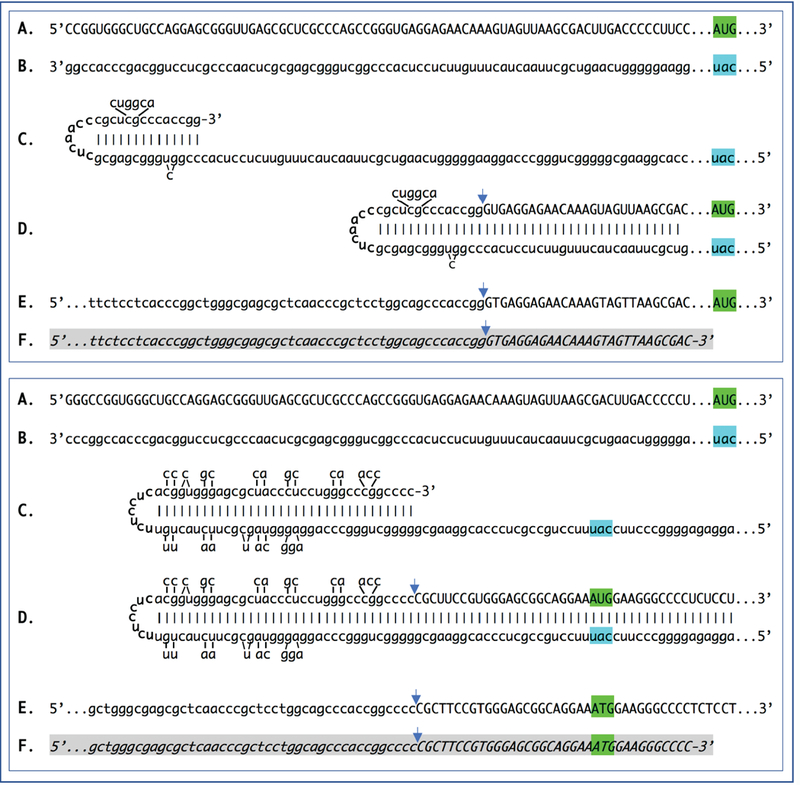
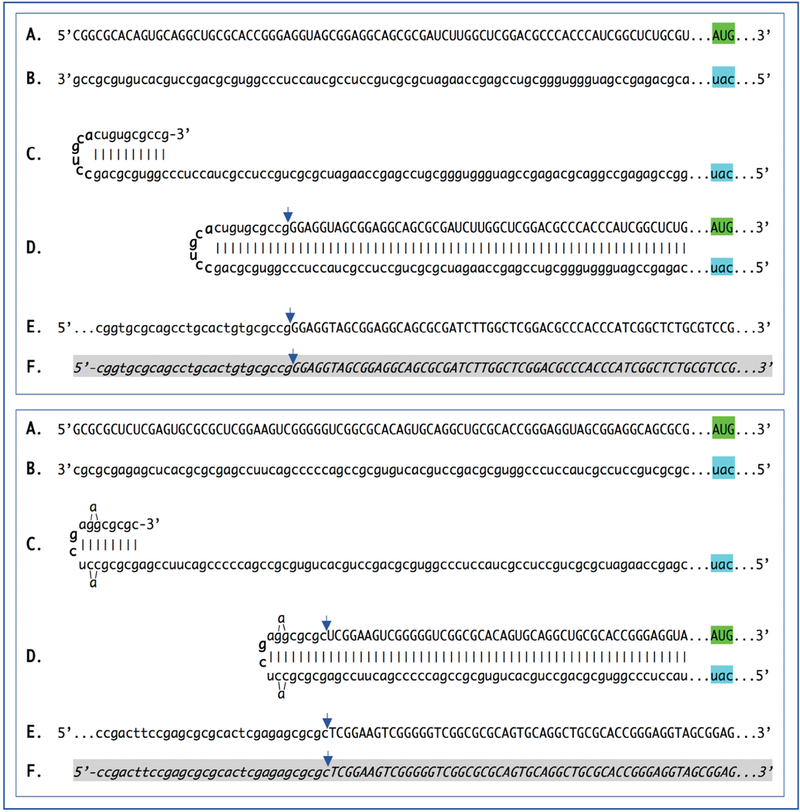
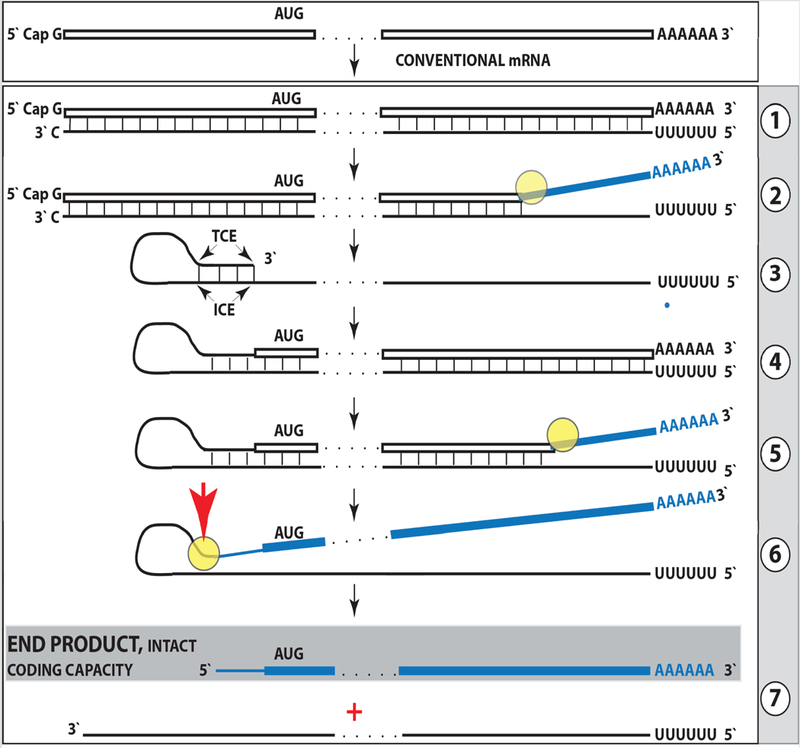
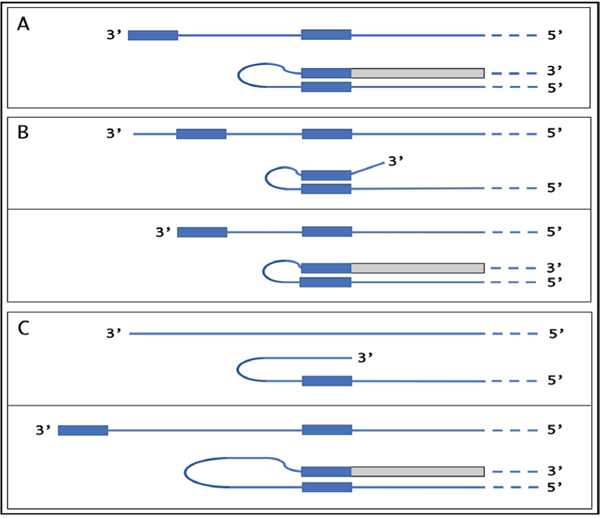
De novo production of RNA on RNA template, a process known as RNA-dependent RNA synthesis, RdRs, and the enzymatic activity conducting it, RNA-dependent RNA polymerase, RdRp, were initially considered to be exclusively virus-specific. Eventually, however, the occurrence of RdRs and the ubiquitous presence of conventional RdRp were demonstrated in numerous eukaryotic organisms. The evidence that the enzymatic machinery capable of RdRs is present in mammalian cells was derived from studies of viruses, such as hepatitis delta virus, HDV, that do not encode RdRp yet undergo a robust RNA replication once inside the mammalian host; thus firmly establishing its occurrence and functionality. Moreover, it became clear that RdRp activity, apparently in a non-conventional form, is constitutively present in most, if not in all, mammalian cells. Because such activity was shown to produce short transcripts, given its apparent involvement in RNA interference phenomena, and because double-stranded RNA is known to trigger cellular responses leading to its degradation, it was generally assumed that its role in mammalian cells is restricted to a regulatory function. However, at the same time, an enzymatic activity capable of generating complete antisense RNA complements of mRNAs was discovered in mammalian cells undergoing terminal differentiation. Moreover, observations of widespread synthesis of antisense RNAs initiating at the 3'poly(A) of mRNAs in human cells suggested an extensive cellular utilization of mammalian RdRp. These results led to the development of a model of RdRp-facilitated and antisense RNA-mediated amplification of mammalian mRNA. Recent detection of the major model-predicted identifiers, chimeric RNA intermediates containing both sense and antisense RNA strands covalently joined in a rigorously predicted and uniquely defined manner, as well as the identification of a putative chimeric RNA end product of this process, validated the proposed model. The results corroborating mammalian RNA-dependent mRNA amplification were obtained in vivo with cells undergoing terminal erythroid differentiation and programmed for only a short survival span. This raises a question of whether mammalian RNA-dependent mRNA amplification is a specialized occurrence limited to extreme circumstances of terminal differentiation or a general physiological phenomenon. The present study addresses this question by testing for the occurrence of RNA-dependent amplification of mRNA encoding extracellular matrix proteins abundantly produced throughout the tissue and organ development and homeostasis, an exceptionally revealing indicator of the range and scope of this phenomenon. We report here the detection of major identifiers of RNA-dependent amplification of mRNA encoding α1, β1, and γ1 chains of laminin in mouse tissues producing large quantities of extracellular matrix proteins. The results obtained warrant reinterpretation of the mechanisms involved in ubiquitous and abundant production and deposition of extracellular matrix proteins, confirm the occurrence of mammalian RNA-dependent mRNA amplification as a new mode of genomic protein-encoding information transfer, and establish it as a general physiological phenomenon.
Keywords: Antisense-strand RNA; Chimeric RNA intermediate; RNA-dependent RNA polymerase; RNA-dependent amplification of mammalian mRNA; Self-priming; Sensestrand RNA.
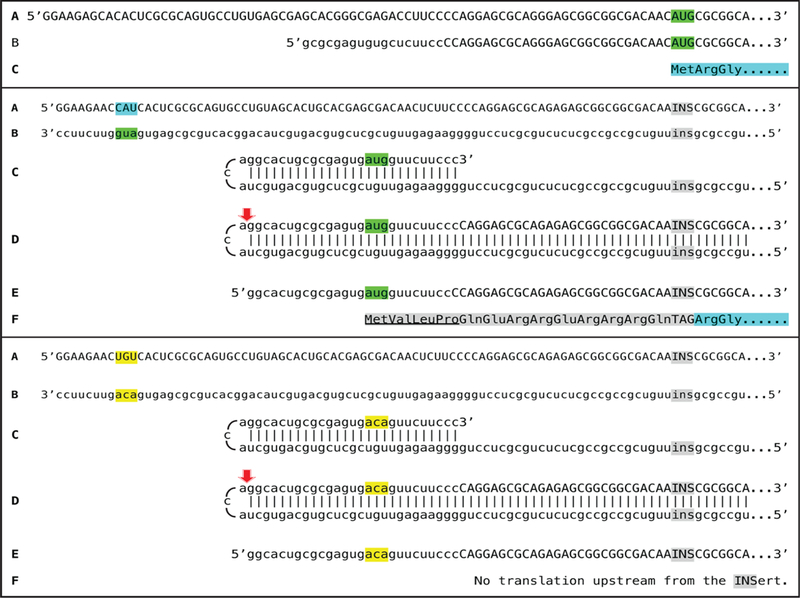
Figures


Similar articles
-
News from Mars: Two-Tier Paradox, Intracellular PCR, Chimeric Junction Shift, Dark Matter mRNA and Other Remarkable Features of Mammalian RNA-Dependent mRNA Amplification. Implications for Alzheimer's Disease, RNA-Based Vaccines and mRNA Therapeutics.Ann Integr Mol Med. 2021;2:131-173. doi: 10.33597/aimm.02-1009. Ann Integr Mol Med. 2021. PMID: 33942036 Free PMC article.
-
Protein-Encoding RNA-to-RNA Information Transfer in Mammalian Cells: Principles of RNA-Dependent mRNA Amplification.Ann Integr Mol Med. 2019;1(1):1002. Ann Integr Mol Med. 2019. PMID: 31535092 Free PMC article.
-
Protein-Encoding RNA to RNA Information Transfer in Mammalian Cells: RNA-dependent mRNA Amplification. Identification of Chimeric RNA Intermediates and Putative RNA End Products.Ann Integr Mol Med. 2019;1(1):23-47. Epub 2019 Aug 22. Ann Integr Mol Med. 2019. PMID: 31656957 Free PMC article.
-
HDV RNA replication: ancient relic or primer?Curr Top Microbiol Immunol. 2006;307:25-45. doi: 10.1007/3-540-29802-9_2. Curr Top Microbiol Immunol. 2006. PMID: 16903219 Review.
-
RNA recombination in hepatitis delta virus: implications regarding the abilities of mammalian RNA polymerases.Virus Res. 2007 Aug;127(2):208-15. doi: 10.1016/j.virusres.2007.01.003. Epub 2007 Feb 12. Virus Res. 2007. PMID: 17296240 Review.
Cited by
-
News from Mars: Two-Tier Paradox, Intracellular PCR, Chimeric Junction Shift, Dark Matter mRNA and Other Remarkable Features of Mammalian RNA-Dependent mRNA Amplification. Implications for Alzheimer's Disease, RNA-Based Vaccines and mRNA Therapeutics.Ann Integr Mol Med. 2021;2:131-173. doi: 10.33597/aimm.02-1009. Ann Integr Mol Med. 2021. PMID: 33942036 Free PMC article.
-
The Amyloid Cascade Hypothesis 2.0: Generalization of the Concept.J Alzheimers Dis Rep. 2023 Jan 19;7(1):21-35. doi: 10.3233/ADR-220079. eCollection 2023. J Alzheimers Dis Rep. 2023. PMID: 36777328 Free PMC article.
-
The Amyloid Cascade Hypothesis 2.0 for Alzheimer's Disease and Aging-Associated Cognitive Decline: From Molecular Basis to Effective Therapy.Int J Mol Sci. 2023 Jul 31;24(15):12246. doi: 10.3390/ijms241512246. Int J Mol Sci. 2023. PMID: 37569624 Free PMC article.
-
Protein-Encoding RNA-to-RNA Information Transfer in Mammalian Cells: Principles of RNA-Dependent mRNA Amplification.Ann Integr Mol Med. 2019;1(1):1002. Ann Integr Mol Med. 2019. PMID: 31535092 Free PMC article.
-
The Amyloid Cascade Hypothesis 2.0: On the Possibility of Once-in-a-Lifetime-Only Treatment for Prevention of Alzheimer's Disease and for Its Potential Cure at Symptomatic Stages.J Alzheimers Dis Rep. 2022 Jul 11;6(1):369-399. doi: 10.3233/ADR-220031. eCollection 2022. J Alzheimers Dis Rep. 2022. PMID: 36072366 Free PMC article.
References
-
- Rits S, Olsen B, Volloch V. RNA-dependent synthesis of mammalian mRNA: Identification of chimeric intermediate and putative end-product. BioRxiv. 2016.
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials