

PDZD8 mediates a Rab7-dependent interaction of the ER with late endosomes and lysosomes
- PMID: 31636202
- PMCID: PMC6842579
- DOI: 10.1073/pnas.1913509116
PDZD8 mediates a Rab7-dependent interaction of the ER with late endosomes and lysosomes
Abstract
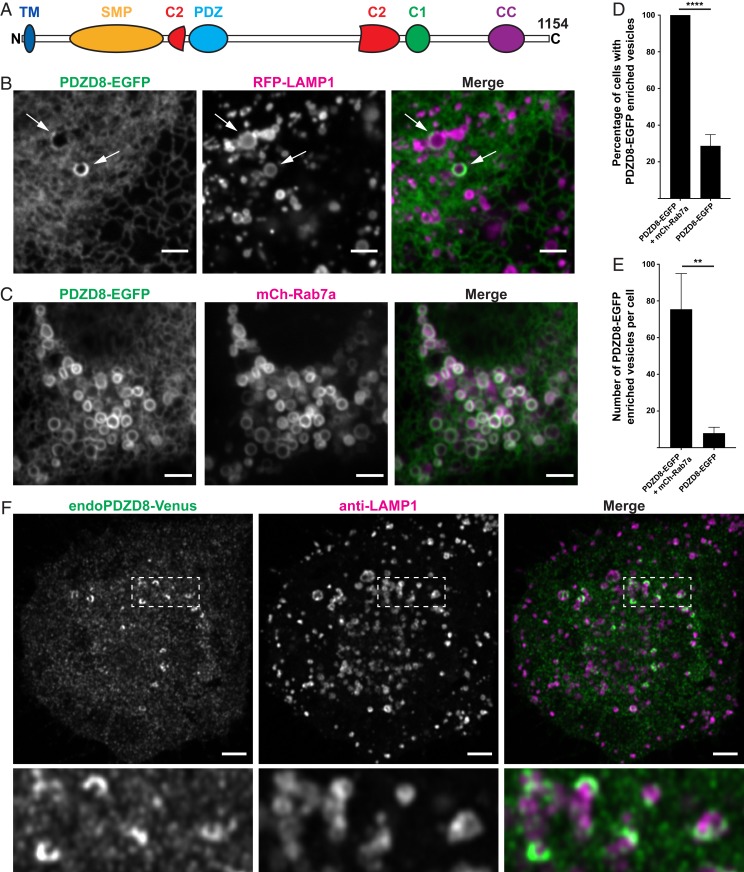
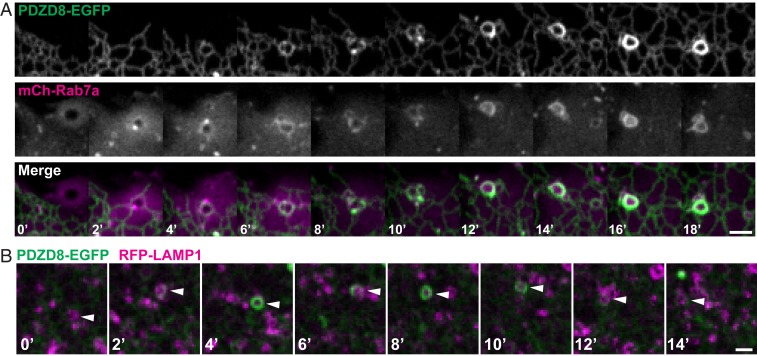
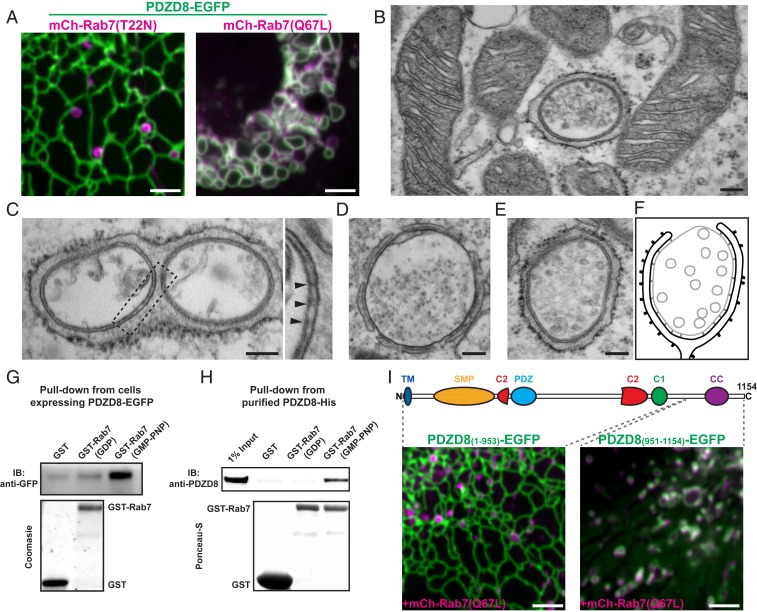
Contacts between the endoplasmic reticulum (ER) and other membranes are hot spots for protein-mediated lipid transport between the 2 adjacent bilayers. Compiling a molecular inventory of lipid transport proteins present at these sites is a premise to the elucidation of their function. Here we show that PDZD8, an intrinsic membrane protein of the ER with a lipid transport module of the SMP domain family, concentrates at contacts between the ER and late endosomes/lysosomes, where it interacts with GTP-Rab7. These findings suggest that PDZD8 may cooperate with other proteins that function at the ER-endo/lysosome interface in coordinating endocytic flow with lipid transport between endocytic membranes and the ER.
Keywords: SMP domain; lipid-transfer protein; membrane contact sites.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Structural basis of human PDZD8-Rab7 interaction for the ER-late endosome tethering.Sci Rep. 2021 Sep 22;11(1):18859. doi: 10.1038/s41598-021-98419-5. Sci Rep. 2021. PMID: 34552186 Free PMC article.
-
PDZD8 interacts with Protrudin and Rab7 at ER-late endosome membrane contact sites associated with mitochondria.Nat Commun. 2020 Jul 20;11(1):3645. doi: 10.1038/s41467-020-17451-7. Nat Commun. 2020. PMID: 32686675 Free PMC article.
-
PDZD8-mediated lipid transfer at contacts between the ER and late endosomes/lysosomes is required for neurite outgrowth.J Cell Sci. 2022 Mar 1;135(5):jcs255026. doi: 10.1242/jcs.255026. Epub 2021 Apr 20. J Cell Sci. 2022. PMID: 33912962
-
Cholesterol transfer at endosomal-organelle membrane contact sites.Curr Opin Lipidol. 2018 Jun;29(3):212-217. doi: 10.1097/MOL.0000000000000506. Curr Opin Lipidol. 2018. PMID: 29629999 Review.
-
Rab GTPase Function in Endosome and Lysosome Biogenesis.Trends Cell Biol. 2018 Nov;28(11):957-970. doi: 10.1016/j.tcb.2018.06.007. Epub 2018 Jul 17. Trends Cell Biol. 2018. PMID: 30025982 Review.
Cited by
-
Tombusviruses Target a Major Crossroad in the Endocytic and Recycling Pathways via Co-opting Rab7 Small GTPase.J Virol. 2021 Oct 13;95(21):e0107621. doi: 10.1128/JVI.01076-21. Epub 2021 Aug 18. J Virol. 2021. PMID: 34406861 Free PMC article.
-
PDZD8 Disruption Causes Cognitive Impairment in Humans, Mice, and Fruit Flies.Biol Psychiatry. 2022 Aug 15;92(4):323-334. doi: 10.1016/j.biopsych.2021.12.017. Epub 2022 Jan 11. Biol Psychiatry. 2022. PMID: 35227461 Free PMC article.
-
Staying in touch with the endocytic network: The importance of contacts for cholesterol transport.Traffic. 2020 May;21(5):354-363. doi: 10.1111/tra.12726. Epub 2020 Mar 31. Traffic. 2020. PMID: 32129938 Free PMC article. Review.
-
AMPK targets PDZD8 to trigger carbon source shift from glucose to glutamine.Cell Res. 2024 Oct;34(10):683-706. doi: 10.1038/s41422-024-00985-6. Epub 2024 Jun 19. Cell Res. 2024. PMID: 38898113 Free PMC article.
-
Lipid Transfer-Dependent Endosome Maturation Mediated by Protrudin and PDZD8 in Neurons.Front Cell Dev Biol. 2020 Dec 15;8:615600. doi: 10.3389/fcell.2020.615600. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33385000 Free PMC article. Review.
References
-
- Wirtz K. W., Phospholipid transfer proteins. Annu. Rev. Biochem. 60, 73–99 (1991). - PubMed
-
- Holthuis J. C. M., Menon A. K., Lipid landscapes and pipelines in membrane homeostasis. Nature 510, 48–57 (2014). - PubMed
-
- Wong L. H., Gatta A. T., Levine T. P., Lipid transfer proteins: The lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 20, 85–101 (2019). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
