

Topological analysis of the gp41 MPER on lipid bilayers relevant to the metastable HIV-1 envelope prefusion state
- PMID: 31624123
- PMCID: PMC6842640
- DOI: 10.1073/pnas.1912427116
Topological analysis of the gp41 MPER on lipid bilayers relevant to the metastable HIV-1 envelope prefusion state
Abstract
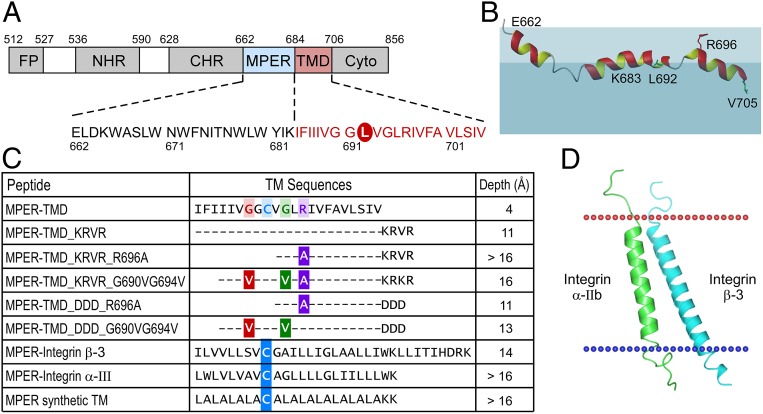
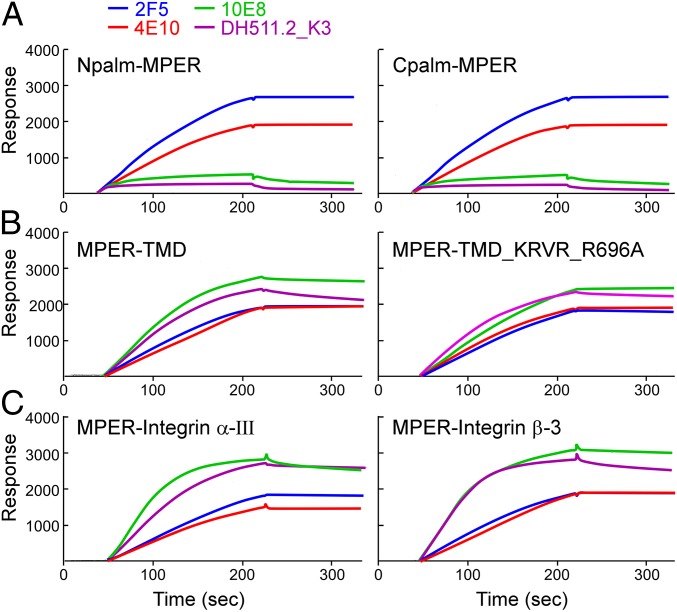
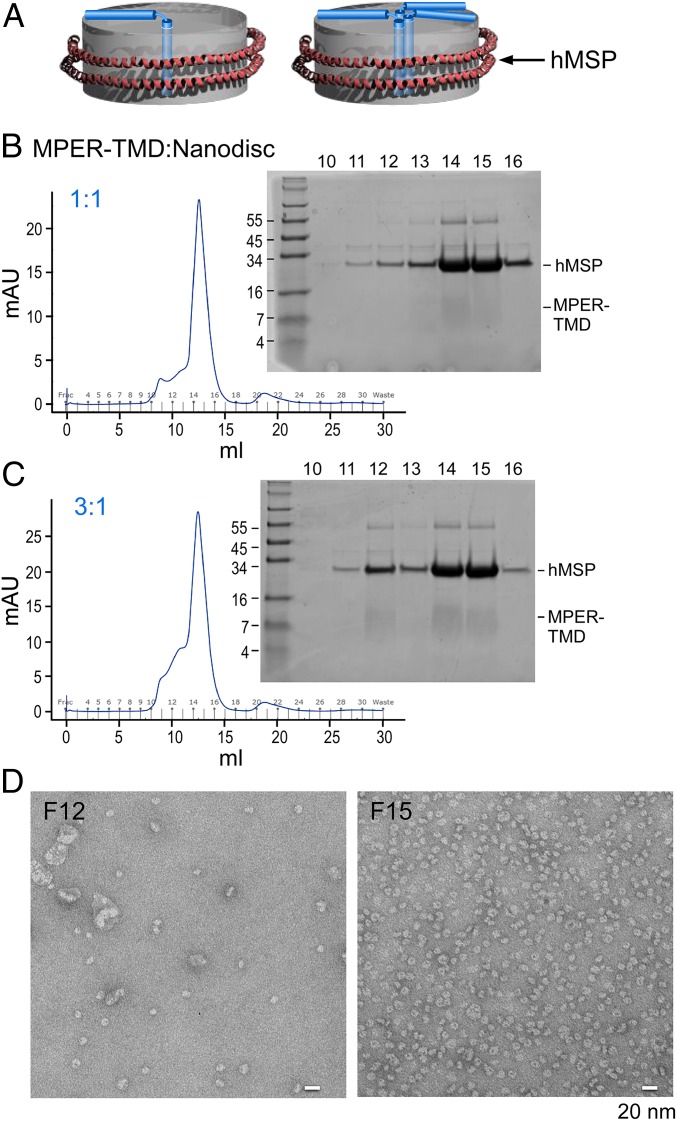
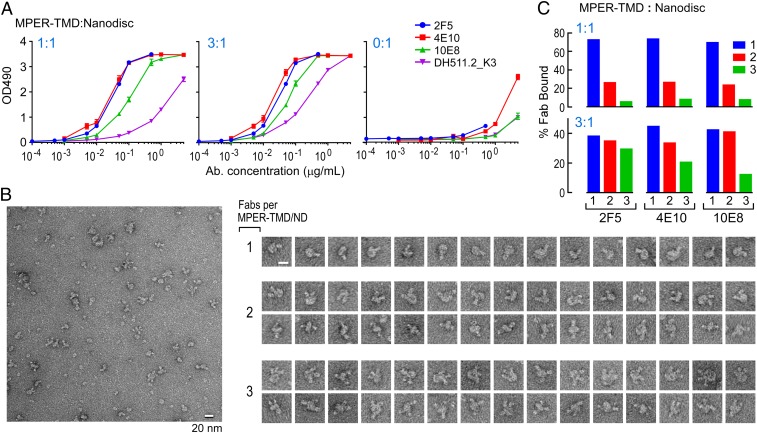
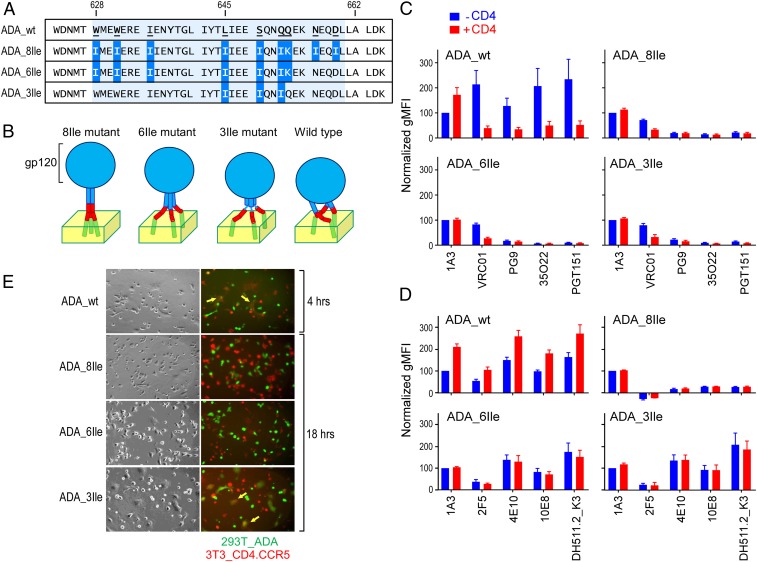
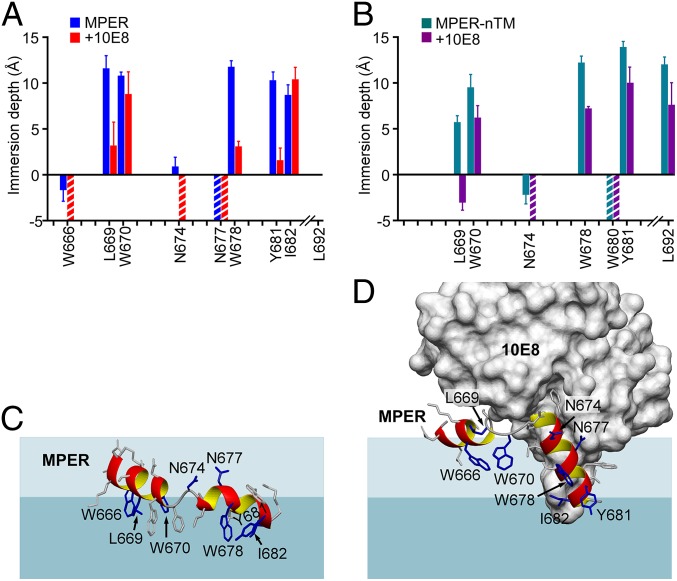
The membrane proximal external region (MPER) of HIV-1 envelope glycoprotein (gp) 41 is an attractive vaccine target for elicitation of broadly neutralizing antibodies (bNAbs) by vaccination. However, current details regarding the quaternary structural organization of the MPER within the native prefusion trimer [(gp120/41)3] are elusive and even contradictory, hindering rational MPER immunogen design. To better understand the structural topology of the MPER on the lipid bilayer, the adjacent transmembrane domain (TMD) was appended (MPER-TMD) and studied. Membrane insertion of the MPER-TMD was sensitive both to the TMD sequence and cytoplasmic residues. Antigen binding of MPER-specific bNAbs, in particular 10E8 and DH511.2_K3, was significantly impacted by the presence of the TMD. Furthermore, MPER-TMD assembly into 10-nm diameter nanodiscs revealed a heterogeneous membrane array comprised largely of monomers and dimers, as enumerated by bNAb Fab binding using single-particle electron microscopy analysis, arguing against preferential trimeric association of native MPER and TMD protein segments. Moreover, introduction of isoleucine mutations in the C-terminal heptad repeat to induce an extended MPER α-helical bundle structure yielded an antigenicity profile of cell surface-arrayed Env variants inconsistent with that found in the native prefusion state. In line with these observations, electron paramagnetic resonance analysis suggested that 10E8 inhibits viral membrane fusion by lifting the MPER N-terminal region out of the viral membrane, mandating the exposure of residues that would be occluded by MPER trimerization. Collectively, our data suggest that the MPER is not a stable trimer, but rather a dynamic segment adapted for structural changes accompanying fusion.
Keywords: Env; HIV-1; MPER; nanodiscs; transmembrane domains.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Exposure of the HIV-1 broadly neutralizing antibody 10E8 MPER epitope on the membrane surface by gp41 transmembrane domain scaffolds.Biochim Biophys Acta Biomembr. 2018 Jun;1860(6):1259-1271. doi: 10.1016/j.bbamem.2018.02.019. Epub 2018 Feb 23. Biochim Biophys Acta Biomembr. 2018. PMID: 29477358
-
Functional Optimization of Broadly Neutralizing HIV-1 Antibody 10E8 by Promotion of Membrane Interactions.J Virol. 2018 Mar 28;92(8):e02249-17. doi: 10.1128/JVI.02249-17. Print 2018 Apr 15. J Virol. 2018. PMID: 29386285 Free PMC article.
-
Mechanism of HIV-1 neutralization by antibodies targeting a membrane-proximal region of gp41.J Virol. 2014 Jan;88(2):1249-58. doi: 10.1128/JVI.02664-13. Epub 2013 Nov 13. J Virol. 2014. PMID: 24227838 Free PMC article.
-
Neutralizing Antibodies Targeting HIV-1 gp41.Viruses. 2020 Oct 23;12(11):1210. doi: 10.3390/v12111210. Viruses. 2020. PMID: 33114242 Free PMC article. Review.
-
Antigp41 membrane proximal external region antibodies and the art of using the membrane for neutralization.Curr Opin HIV AIDS. 2017 May;12(3):250-256. doi: 10.1097/COH.0000000000000364. Curr Opin HIV AIDS. 2017. PMID: 28422789 Review.
Cited by
-
Global Increases in Human Immunodeficiency Virus Neutralization Sensitivity Due to Alterations in the Membrane-Proximal External Region of the Envelope Glycoprotein Can Be Minimized by Distant State 1-Stabilizing Changes.J Virol. 2022 Apr 13;96(7):e0187821. doi: 10.1128/jvi.01878-21. Epub 2022 Mar 15. J Virol. 2022. PMID: 35289647 Free PMC article.
-
Priming with DNA Expressing Trimeric HIV V1V2 Alters the Immune Hierarchy Favoring the Development of V2-Specific Antibodies in Rhesus Macaques.J Virol. 2020 Dec 22;95(2):e01193-20. doi: 10.1128/JVI.01193-20. Print 2020 Dec 22. J Virol. 2020. PMID: 33087466 Free PMC article.
-
DNA-nanostructure-templated assembly of planar and curved lipid-bilayer membranes.Front Chem. 2023 Feb 8;10:1047874. doi: 10.3389/fchem.2022.1047874. eCollection 2022. Front Chem. 2023. PMID: 36844038 Free PMC article. Review.
-
Metastable HIV-1 Surface Protein Env Sensitizes Cell Membranes to Transformation and Poration by Dual-Acting Virucidal Entry Inhibitors.Biochemistry. 2020 Feb 18;59(6):818-828. doi: 10.1021/acs.biochem.9b01008. Epub 2020 Jan 28. Biochemistry. 2020. PMID: 31942789 Free PMC article.
-
Molecular recognition of a membrane-anchored HIV-1 pan-neutralizing epitope.Commun Biol. 2022 Nov 18;5(1):1265. doi: 10.1038/s42003-022-04219-6. Commun Biol. 2022. PMID: 36400835 Free PMC article.
References
-
- Deng H., et al. , Identification of a major co-receptor for primary isolates of HIV-1. Nature 381, 661–666 (1996). - PubMed
-
- Feng Y., Broder C. C., Kennedy P. E., Berger E. A., HIV-1 entry cofactor: Functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science 272, 872–877 (1996). - PubMed
-
- Maddon P. J., et al. , The T4 gene encodes the AIDS virus receptor and is expressed in the immune system and the brain. Cell 47, 333–348 (1986). - PubMed
-
- Chan D. C., Fass D., Berger J. M., Kim P. S., Core structure of gp41 from the HIV envelope glycoprotein. Cell 89, 263–273 (1997). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
