

BAX, BAK, and BOK: A Coming of Age for the BCL-2 Family Effector Proteins
- PMID: 31570337
- PMCID: PMC7111251
- DOI: 10.1101/cshperspect.a036319
BAX, BAK, and BOK: A Coming of Age for the BCL-2 Family Effector Proteins
Abstract
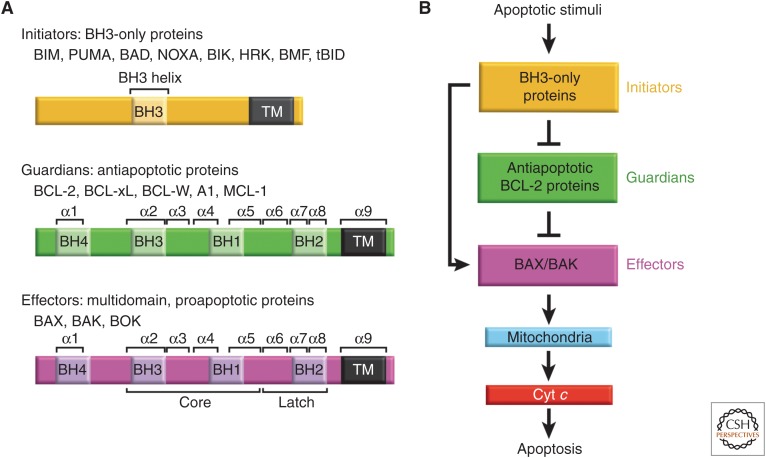
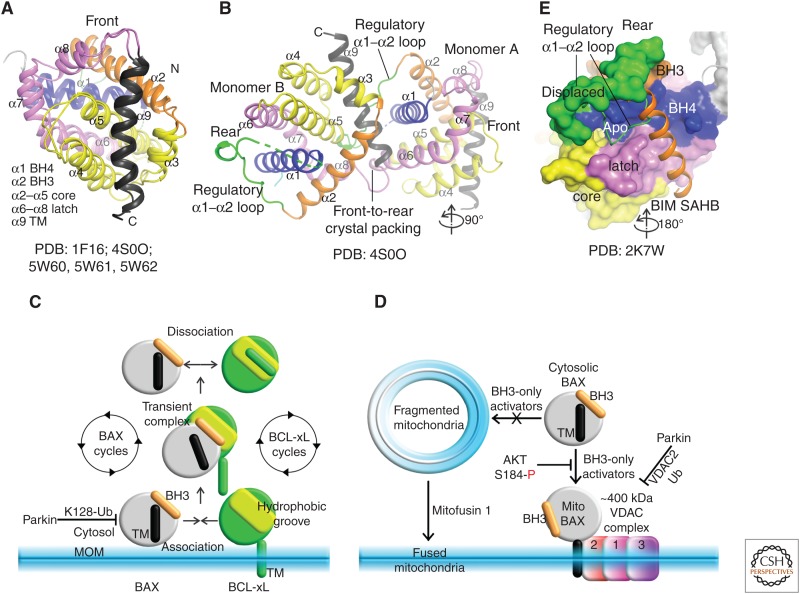
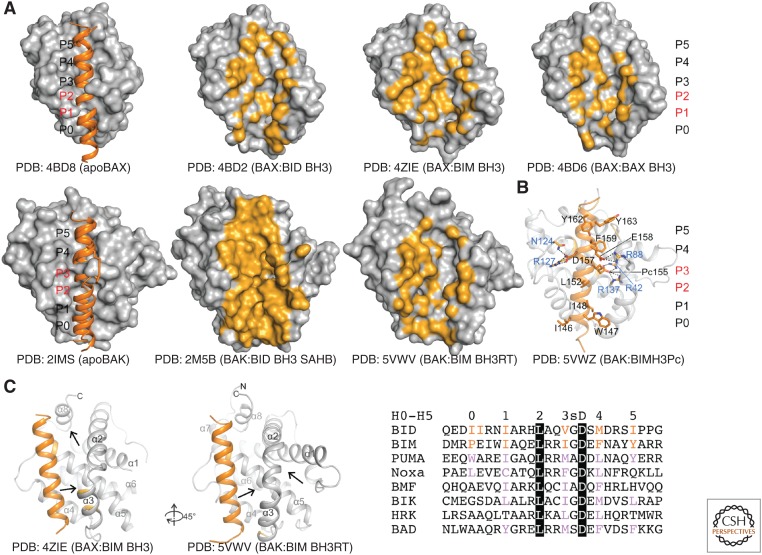
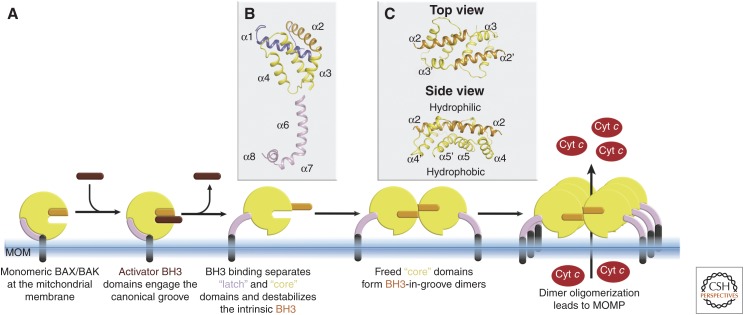
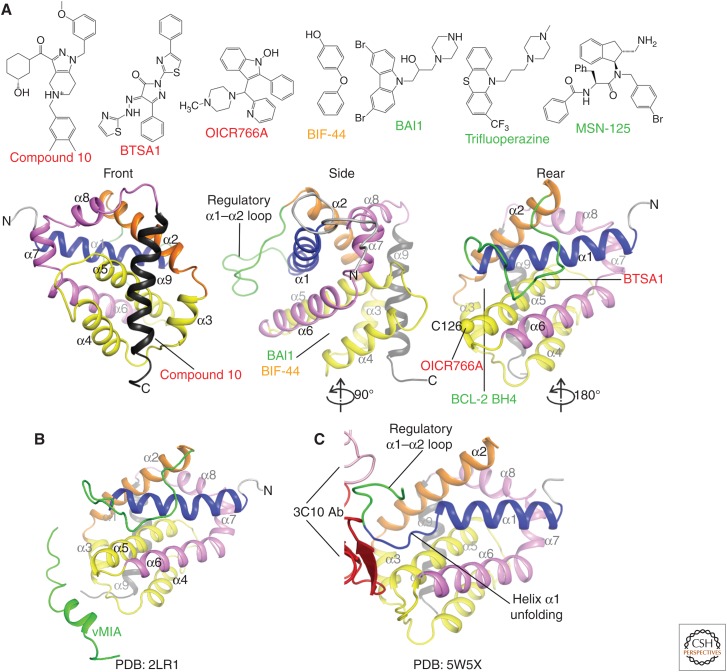
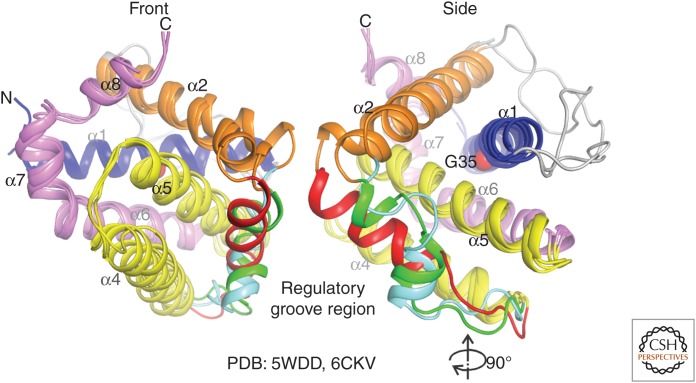
The BCL-2 family of proteins control a key checkpoint in apoptosis, that of mitochondrial outer membrane permeabilization or, simply, mitochondrial poration. The family consists of three subgroups: BH3-only initiators that respond to apoptotic stimuli; antiapoptotic guardians that protect against cell death; and the membrane permeabilizing effectors BAX, BAK, and BOK. On activation, effector proteins are converted from inert monomers into membrane permeabilizing oligomers. For many years, this process has been poorly understood at the molecular level, but a number of recent advances have provided important insights. We review the regulation of these effectors, their activation, subsequent conformational changes, and the ensuing oligomerization events that enable mitochondrial poration, which initiates apoptosis through release of key signaling factors such as cytochrome c We highlight the mysteries that remain in understanding these important proteins in an endeavor to provide a comprehensive picture of where the field currently sits and where it is moving toward.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
Similar articles
-
Apoptotic mitochondrial poration by a growing list of pore-forming BCL-2 family proteins.Bioessays. 2023 Mar;45(3):e2200221. doi: 10.1002/bies.202200221. Epub 2023 Jan 17. Bioessays. 2023. PMID: 36650950 Free PMC article.
-
Contribution of BH3-domain and Transmembrane-domain to the Activity and Interaction of the Pore-forming Bcl-2 Proteins Bok, Bak, and Bax.Sci Rep. 2018 Aug 20;8(1):12434. doi: 10.1038/s41598-018-30603-6. Sci Rep. 2018. PMID: 30127460 Free PMC article.
-
Visualization of BOK pores independent of BAX and BAK reveals a similar mechanism with differing regulation.Cell Death Differ. 2023 Mar;30(3):731-741. doi: 10.1038/s41418-022-01078-w. Epub 2022 Oct 26. Cell Death Differ. 2023. PMID: 36289446 Free PMC article.
-
Discoveries and controversies in BCL-2 protein-mediated apoptosis.FEBS J. 2016 Jul;283(14):2690-700. doi: 10.1111/febs.13527. Epub 2015 Oct 27. FEBS J. 2016. PMID: 26411300 Review.
-
BH3-only proteins trigger cytochrome c release, but how?Arch Biochem Biophys. 2007 Jun 15;462(2):150-5. doi: 10.1016/j.abb.2006.12.022. Epub 2007 Jan 9. Arch Biochem Biophys. 2007. PMID: 17306210 Review.
Cited by
-
CYP2J2-derived epoxyeicosatrienoic acids protect against doxorubicin-induced cardiotoxicity by reducing oxidative stress and apoptosis via activation of the AMPK pathway.Heliyon. 2023 Dec 9;10(1):e23526. doi: 10.1016/j.heliyon.2023.e23526. eCollection 2024 Jan 15. Heliyon. 2023. PMID: 38173517 Free PMC article.
-
The mitochondrial anti-apoptotic dependencies of hematologic malignancies: from disease biology to advances in precision medicine.Haematologica. 2022 Apr 1;107(4):790-802. doi: 10.3324/haematol.2021.280201. Haematologica. 2022. PMID: 35045693 Free PMC article. Review.
-
Trehalose Ameliorates Diabetic Cardiomyopathy: Role of the PK2/PKR Pathway.Oxid Med Cell Longev. 2021 Dec 21;2021:6779559. doi: 10.1155/2021/6779559. eCollection 2021. Oxid Med Cell Longev. 2021. PMID: 34970418 Free PMC article.
-
A novel inhibitory BAK antibody enables assessment of non-activated BAK in cancer cells.Cell Death Differ. 2024 Jun;31(6):711-721. doi: 10.1038/s41418-024-01289-3. Epub 2024 Apr 6. Cell Death Differ. 2024. PMID: 38582955 Free PMC article.
-
The Mysteries around the BCL-2 Family Member BOK.Biomolecules. 2020 Dec 4;10(12):1638. doi: 10.3390/biom10121638. Biomolecules. 2020. PMID: 33291826 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials