

Tanycytic TSPO inhibition induces lipophagy to regulate lipid metabolism and improve energy balance
- PMID: 31469345
- PMCID: PMC7469491
- DOI: 10.1080/15548627.2019.1659616
Tanycytic TSPO inhibition induces lipophagy to regulate lipid metabolism and improve energy balance
Abstract
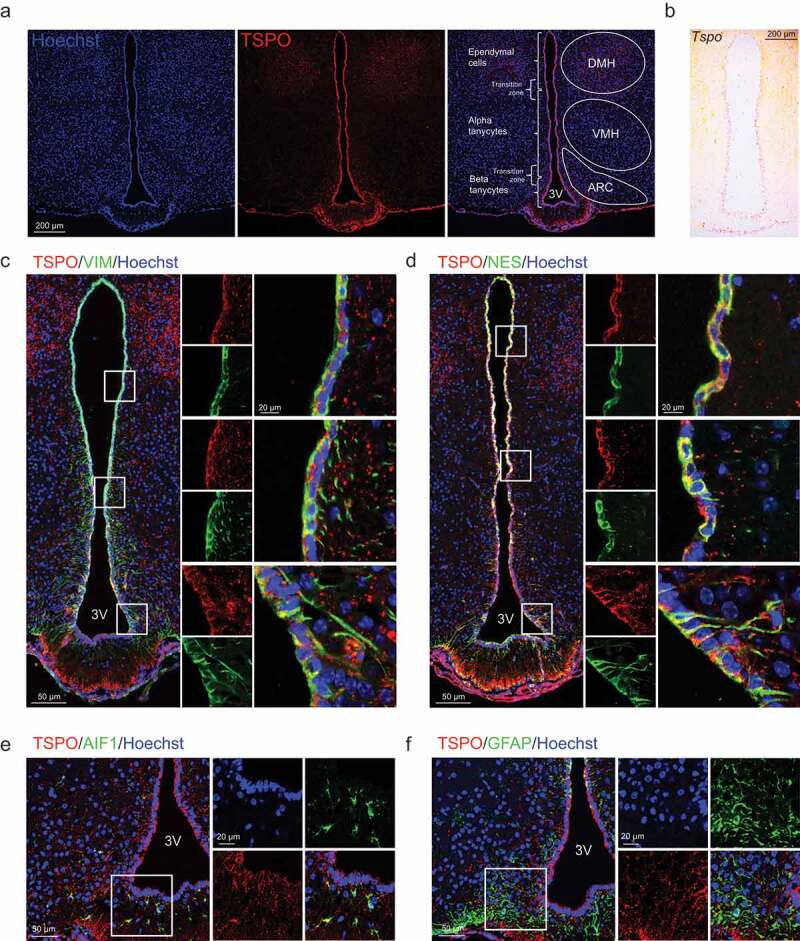
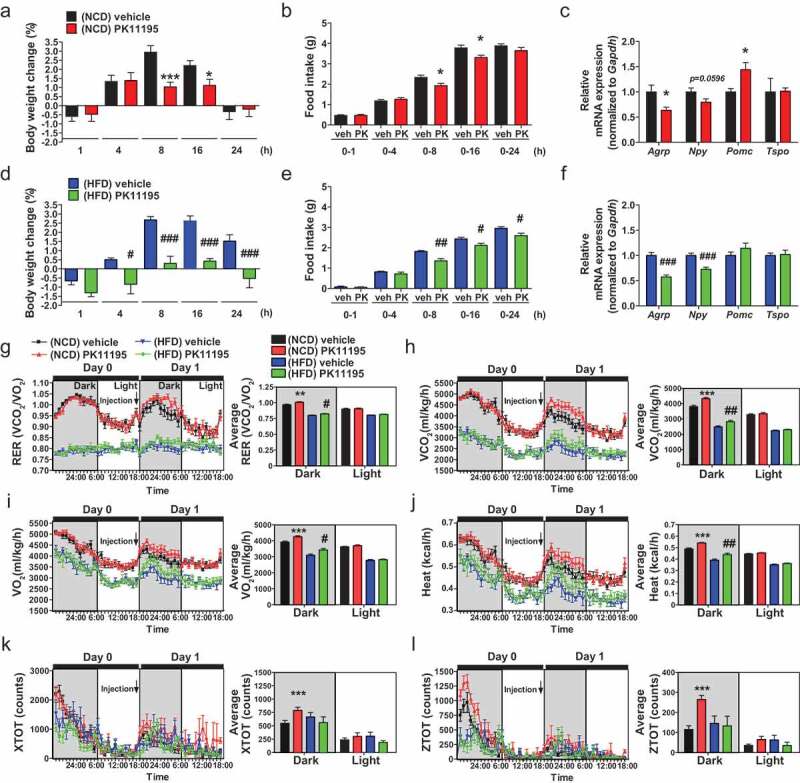
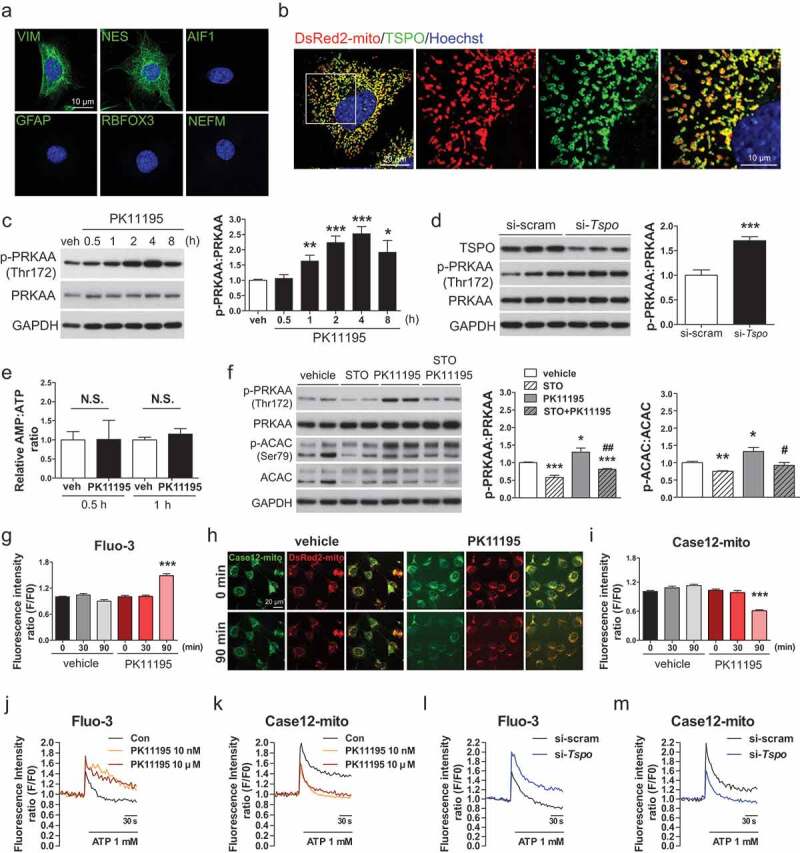
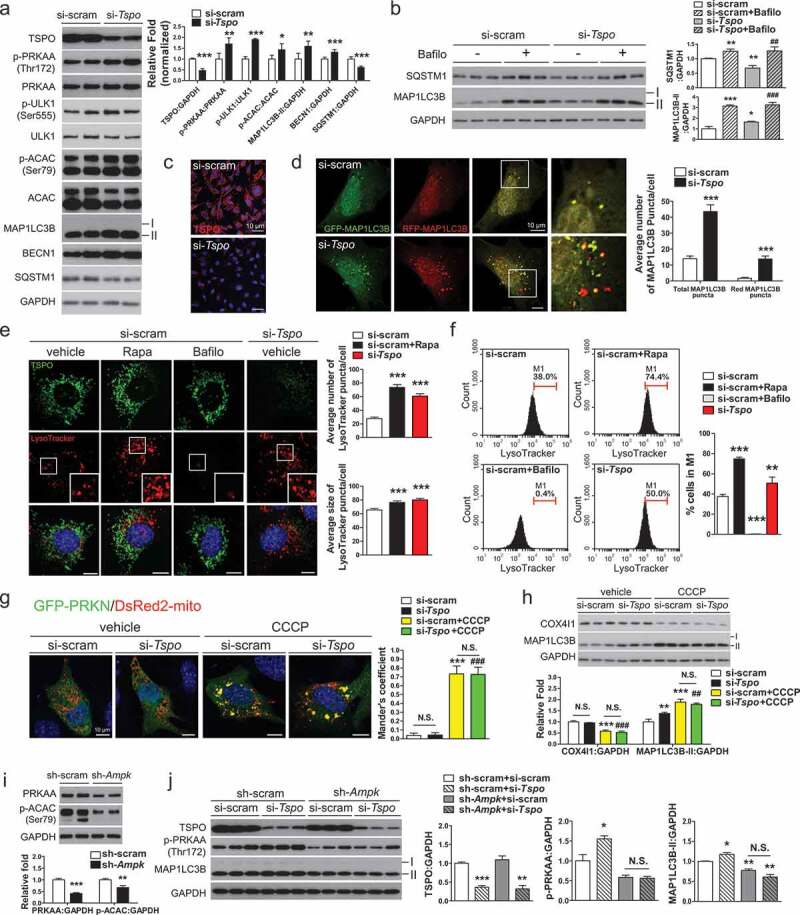
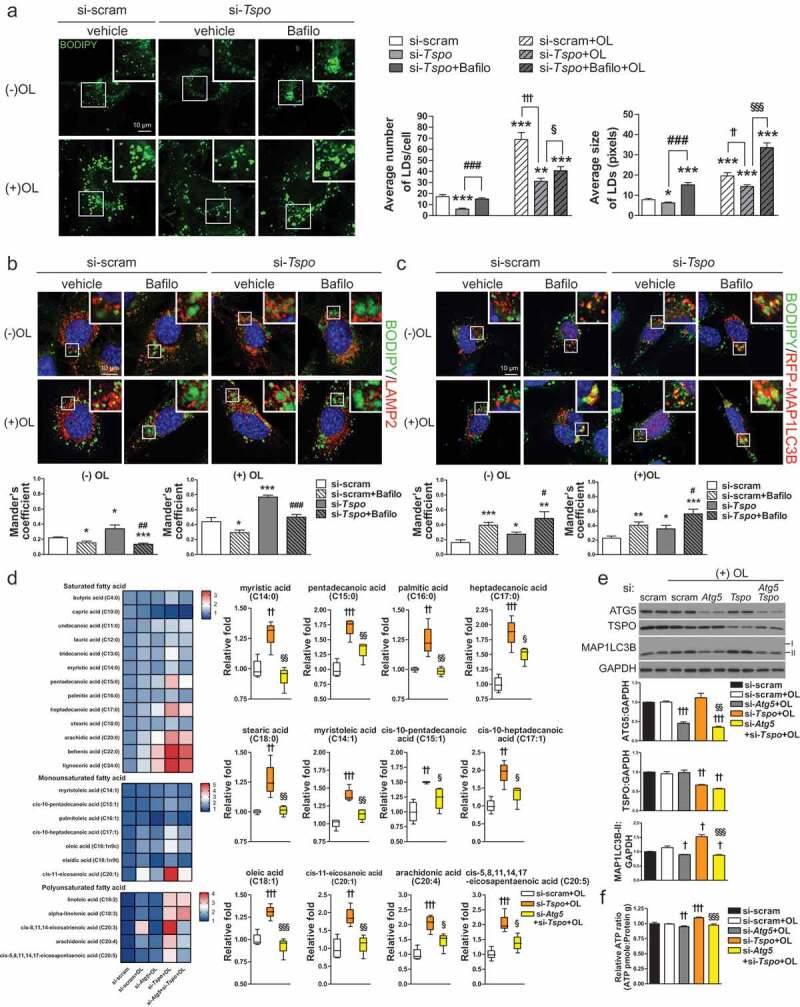
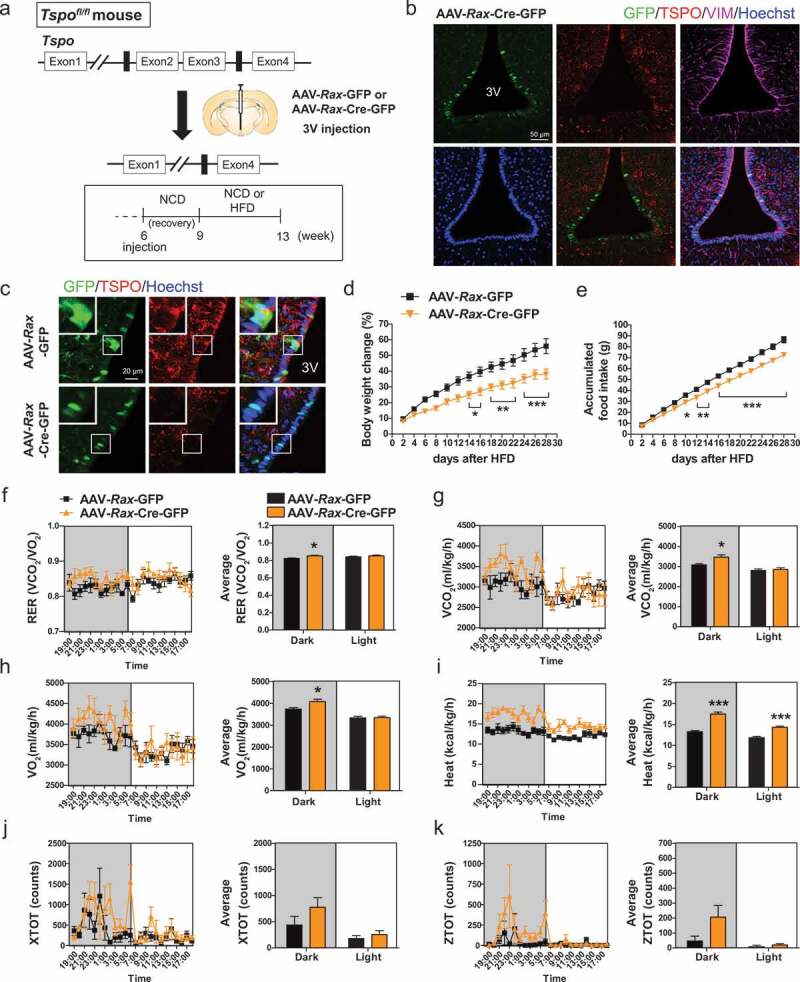
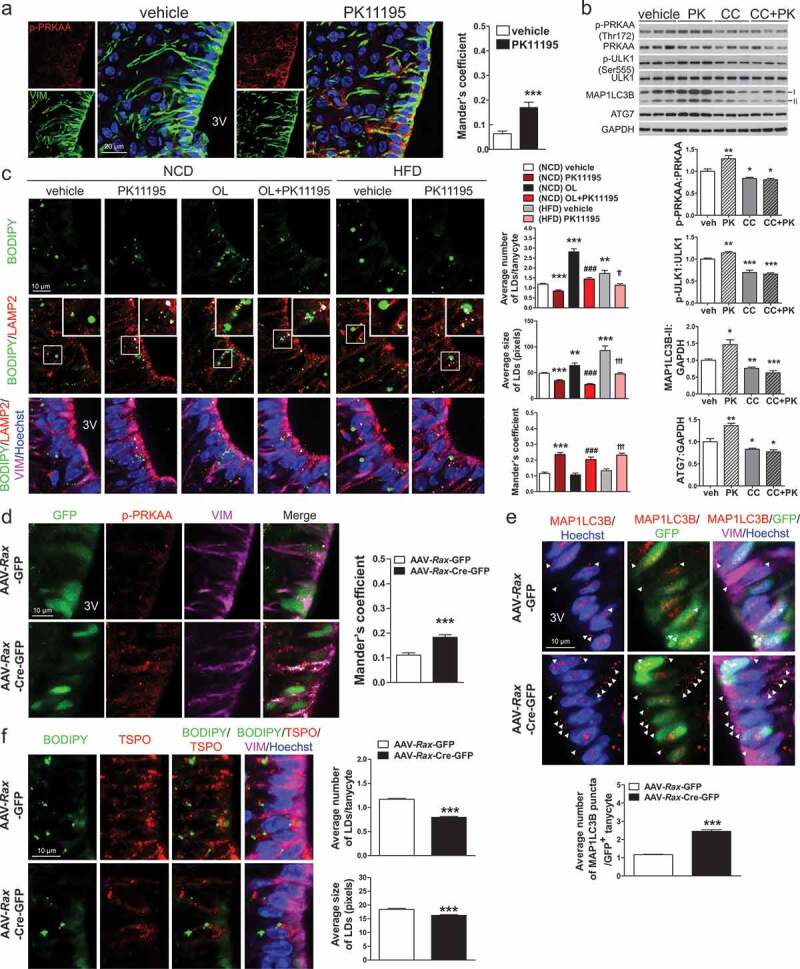
Hypothalamic glial cells named tanycytes, which line the 3rd ventricle (3V), are components of the hypothalamic network that regulates a diverse array of metabolic functions for energy homeostasis. Herein, we report that TSPO (translocator protein), an outer mitochondrial protein, is highly enriched in tanycytes and regulates homeostatic responses to nutrient excess as a potential target for an effective intervention in obesity. Administration of a TSPO ligand, PK11195, into the 3V, and tanycyte-specific deletion of Tspo reduced food intake and elevated energy expenditure, leading to negative energy balance in a high-fat diet challenge. Ablation of tanycytic Tspo elicited AMPK-dependent lipophagy, breaking down lipid droplets into free fatty acids, thereby elevating ATP in a lipid stimulus. Our findings suggest that tanycytic TSPO affects systemic energy balance through macroautophagy/autophagy-regulated lipid metabolism, and highlight the physiological significance of TSPO in hypothalamic lipid sensing and bioenergetics in response to overnutrition.
Abbreviations: 3V: 3rd ventricle; ACAC: acetyl-Coenzyme A carboxylase; AGRP: agouti related neuropeptide; AIF1/IBA1: allograft inflammatory factor 1; AMPK: AMP-activated protein kinase; ARC: arcuate nucleus; Atg: autophagy related; Bafilo: bafilomycin A1; CAMKK2: calcium/calmodulin-dependent protein kinase kinase 2, beta; CCCP: carbonyl cyanide m-chlorophenylhydrazone; CNS: central nervous system; COX4I1: cytochrome c oxidase subunit 4I1; FFA: free fatty acid; GFAP: glial fibrillary acidic protein; HFD: high-fat diet; ICV: intracerebroventricular; LAMP2: lysosomal-associated membrane protein 2; LD: lipid droplet; MAP1LC3B/LC3B: microtubule-associated protein 1 light chain 3 beta; MBH: mediobasal hypothalamus; ME: median eminence; MEF: mouse embryonic fibroblast; NCD: normal chow diet; NEFM/NFM: neurofilament medium; NPY: neuropeptide Y; OL: oleic acid; POMC: pro-opiomelanocortin-alpha; PRKN/Parkin: parkin RBR E3 ubiquitin protein ligase; Rax: retina and anterior neural fold homeobox; RBFOX3/NeuN: RNA binding protein, fox-1 homolog (C. elegans) 3; RER: respiratory exchange ratio; siRNA: small interfering RNA; SQSTM1: sequestosome 1; TG: triglyceride; TSPO: translocator protein; ULK1: unc-51 like kinase 1; VCO2: carbon dioxide production; VMH: ventromedial hypothalamus; VO2: oxygen consumption.
Keywords: AMPK; TSPO; autophagy; energy balance; energy expenditure; food intake; hypothalamus; lipid metabolism; lipophagy; tanycyte.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures
Similar articles
-
The ménage à trois of autophagy, lipid droplets and liver disease.Autophagy. 2022 Jan;18(1):50-72. doi: 10.1080/15548627.2021.1895658. Epub 2021 Apr 2. Autophagy. 2022. PMID: 33794741 Free PMC article. Review.
-
Mitochondrial respiratory chain deficiency inhibits lysosomal hydrolysis.Autophagy. 2019 Sep;15(9):1572-1591. doi: 10.1080/15548627.2019.1586256. Epub 2019 Mar 27. Autophagy. 2019. PMID: 30917721 Free PMC article.
-
Impaired lipophagy induced-microglial lipid droplets accumulation contributes to the buildup of TREM1 in diabetes-associated cognitive impairment.Autophagy. 2023 Oct;19(10):2639-2656. doi: 10.1080/15548627.2023.2213984. Epub 2023 May 19. Autophagy. 2023. PMID: 37204119 Free PMC article.
-
Muscle-generated BDNF (brain derived neurotrophic factor) maintains mitochondrial quality control in female mice.Autophagy. 2022 Jun;18(6):1367-1384. doi: 10.1080/15548627.2021.1985257. Epub 2021 Oct 25. Autophagy. 2022. PMID: 34689722 Free PMC article.
-
Tanycytes and hypothalamic control of energy metabolism.Glia. 2018 Jun;66(6):1176-1184. doi: 10.1002/glia.23303. Epub 2018 Feb 7. Glia. 2018. PMID: 29411421 Review.
Cited by
-
Doublecortin-like knockdown in mice attenuates obesity by stimulating energy expenditure in adipose tissue.Sci Rep. 2024 Aug 22;14(1):19517. doi: 10.1038/s41598-024-70639-5. Sci Rep. 2024. PMID: 39174821 Free PMC article.
-
Palmitate reduces starvation-induced ER stress by inhibiting ER-phagy in hypothalamic cells.Mol Brain. 2021 Apr 6;14(1):65. doi: 10.1186/s13041-021-00777-8. Mol Brain. 2021. PMID: 33823883 Free PMC article.
-
TSPO: an emerging role in appetite for a therapeutically promising biomarker.Open Biol. 2021 Aug;11(8):210173. doi: 10.1098/rsob.210173. Epub 2021 Aug 4. Open Biol. 2021. PMID: 34343461 Free PMC article. Review.
-
Inhibition of ACAT as a Therapeutic Target for Alzheimer's Disease Is Independent of ApoE4 Lipidation.Neurotherapeutics. 2023 Jul;20(4):1120-1137. doi: 10.1007/s13311-023-01375-3. Epub 2023 May 8. Neurotherapeutics. 2023. PMID: 37157042 Free PMC article.
-
The Translocator Protein (TSPO) in Mitochondrial Bioenergetics and Immune Processes.Cells. 2020 Feb 24;9(2):512. doi: 10.3390/cells9020512. Cells. 2020. PMID: 32102369 Free PMC article. Review.
References
-
- Schwartz MW, Woods SC, Porte D Jr.. et al. Central nervous system control of food intake. Nature. 2000. April 06;404(6778):661–671. PubMed PMID: 10766253. - PubMed
-
- Waterson MJ, Horvath TL.. Neuronal regulation of energy homeostasis: beyond the hypothalamus and feeding. Cell Metab. 2015. December 1;22(6):962–970. PubMed PMID: 26603190. - PubMed
-
- Ebling FJP, Lewis JE. Tanycytes and hypothalamic control of energy metabolism. Glia. 2018. June;66(6):1176–1184. PubMed PMID: 29411421. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous