

A wave of monocytes is recruited to replenish the long-term Langerhans cell network after immune injury
- PMID: 31444235
- PMCID: PMC6894529
- DOI: 10.1126/sciimmunol.aax8704
A wave of monocytes is recruited to replenish the long-term Langerhans cell network after immune injury
Abstract
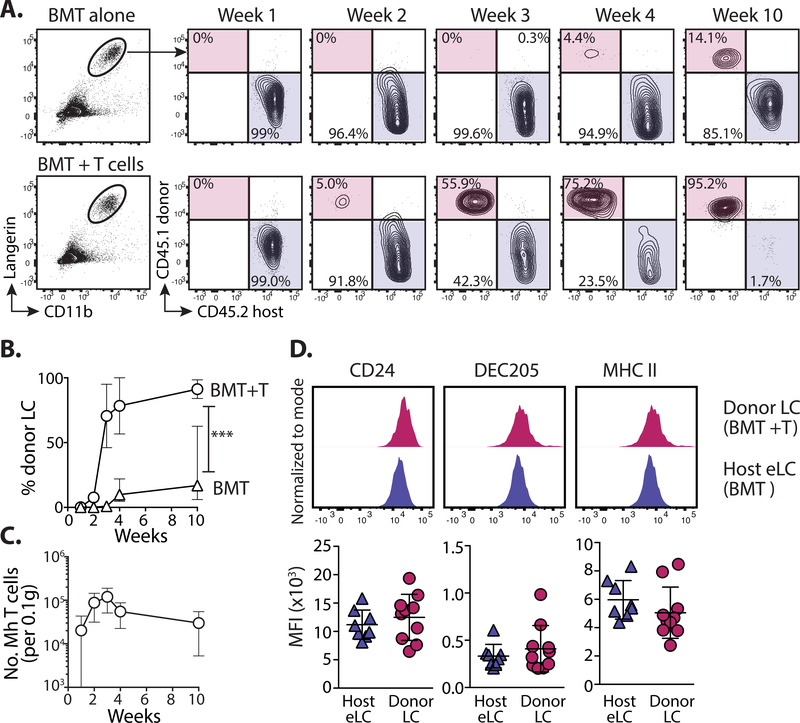
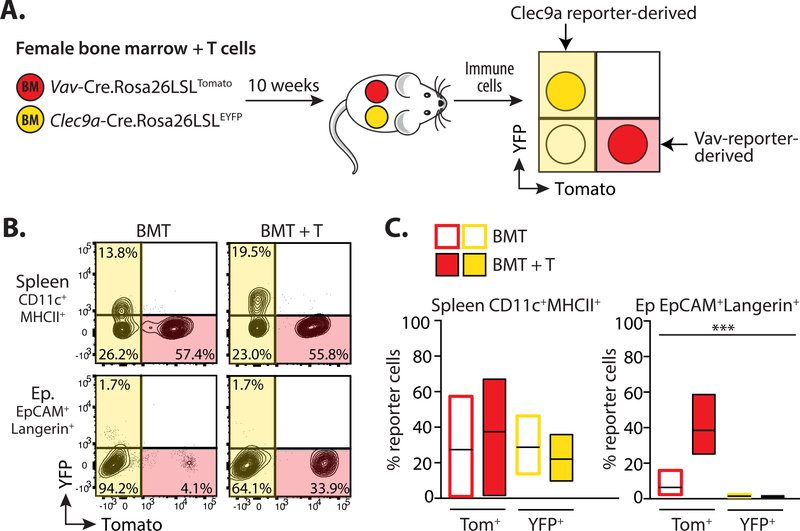
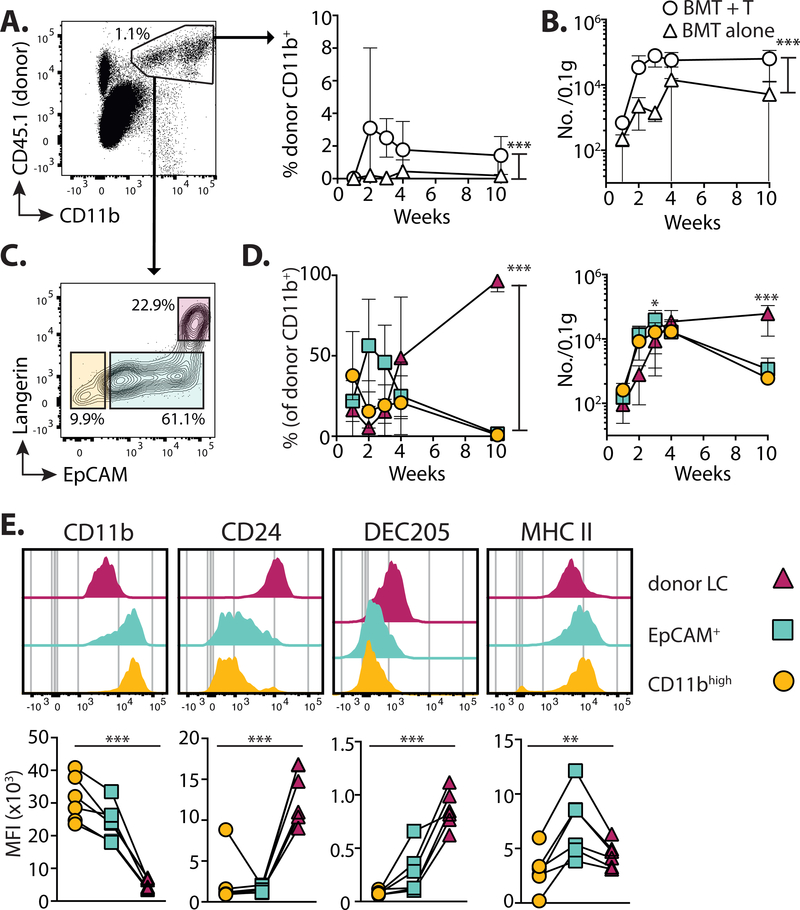
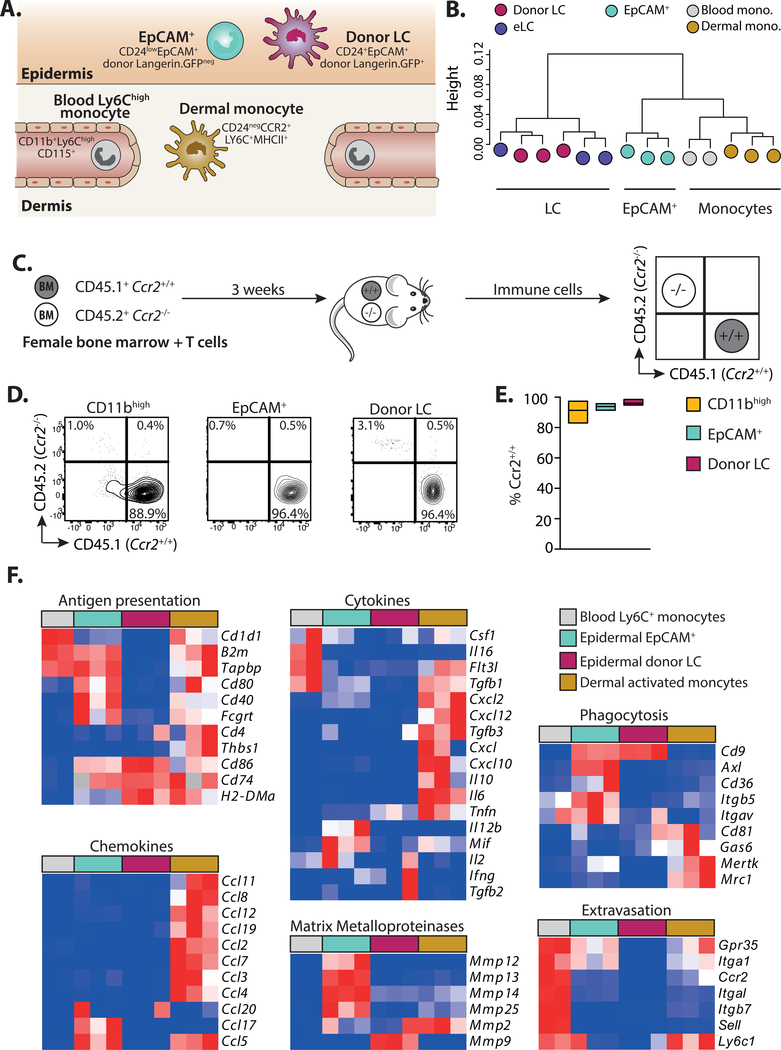
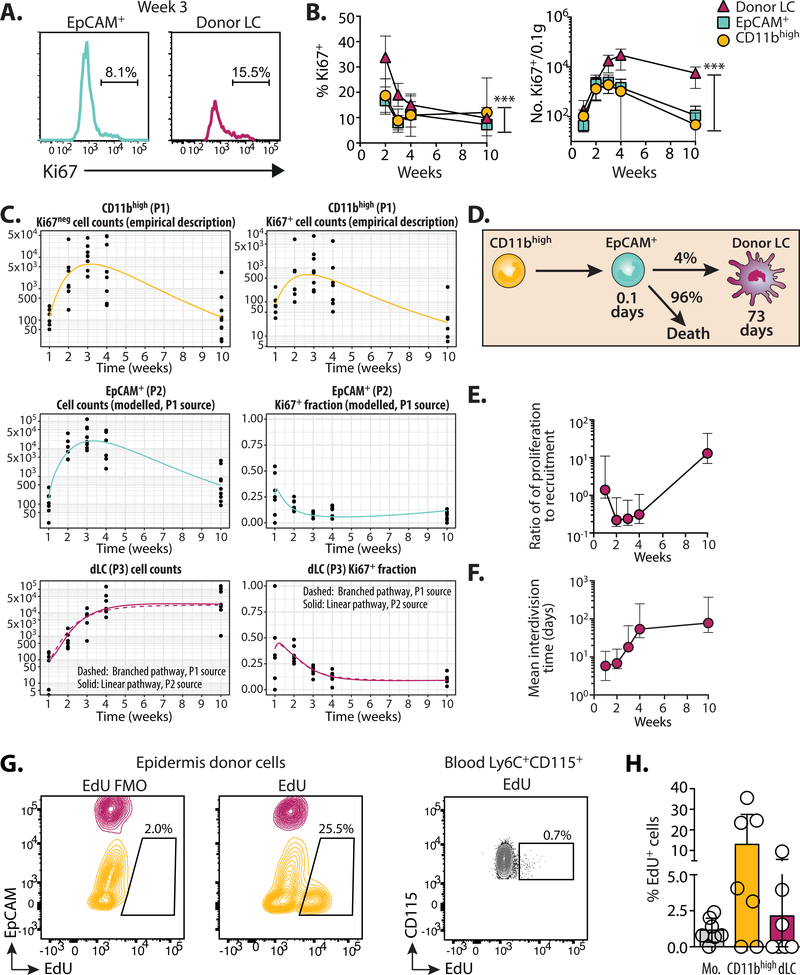
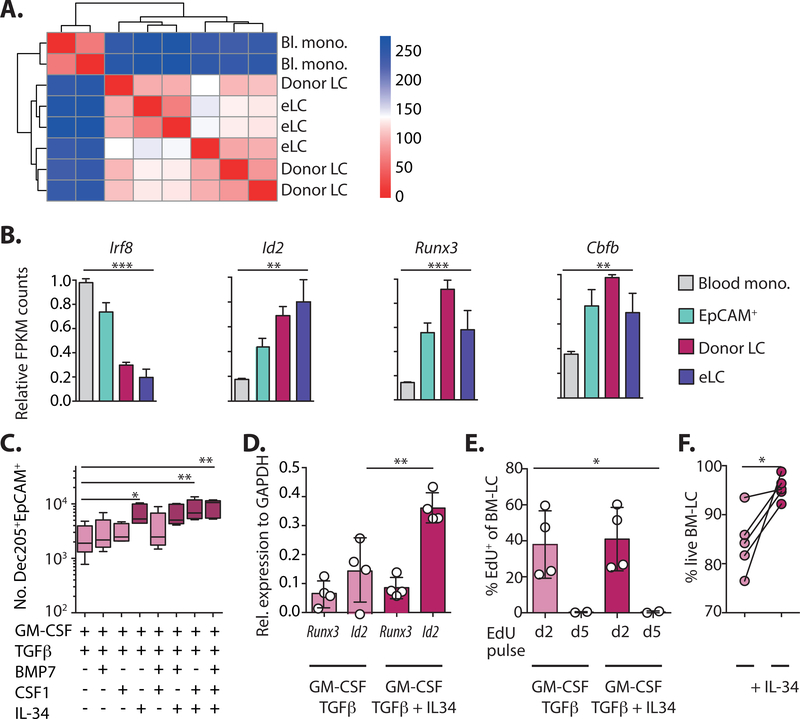
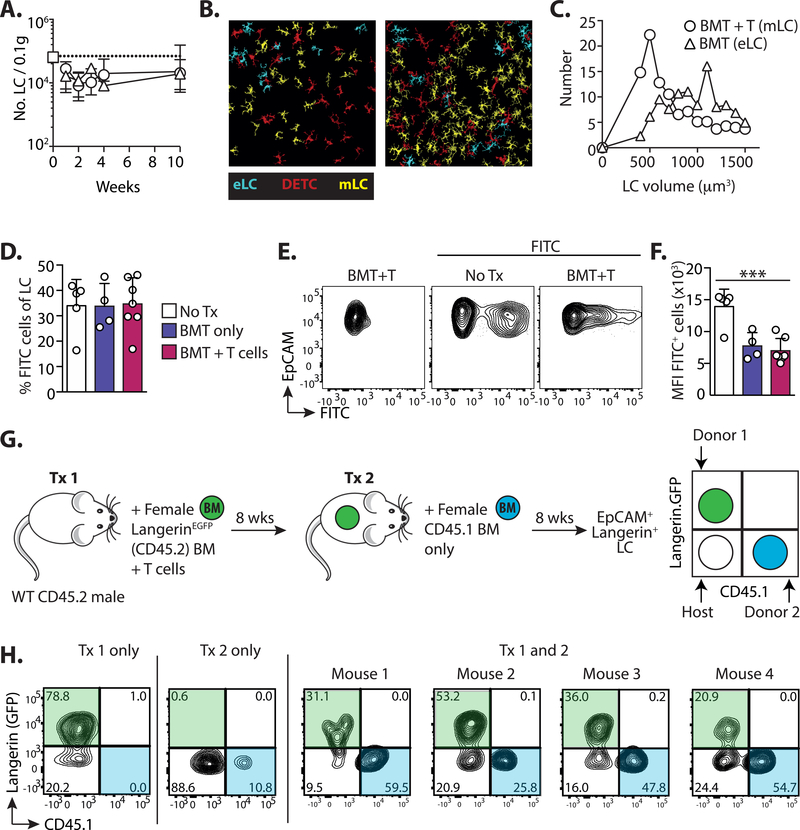
A dense population of embryo-derived Langerhans cells (eLCs) is maintained within the sealed epidermis without contribution from circulating cells. When this network is perturbed by transient exposure to ultraviolet light, short-term LCs are temporarily reconstituted from an initial wave of monocytes but thought to be superseded by more permanent repopulation with undefined LC precursors. However, the extent to which this process is relevant to immunopathological processes that damage LC population integrity is not known. Using a model of allogeneic hematopoietic stem cell transplantation, where alloreactive T cells directly target eLCs, we have asked whether and how the original LC network is ultimately restored. We find that donor monocytes, but not dendritic cells, are the precursors of long-term LCs in this context. Destruction of eLCs leads to recruitment of a wave of monocytes that engraft in the epidermis and undergo a sequential pathway of differentiation via transcriptionally distinct EpCAM+ precursors. Monocyte-derived LCs acquire the capacity of self-renewal, and proliferation in the epidermis matched that of steady-state eLCs. However, we identified a bottleneck in the differentiation and survival of epidermal monocytes, which, together with the slow rate of renewal of mature LCs, limits repair of the network. Furthermore, replenishment of the LC network leads to constitutive entry of cells into the epidermal compartment. Thus, immune injury triggers functional adaptation of mechanisms used to maintain tissue-resident macrophages at other sites, but this process is highly inefficient in the skin.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Similar articles
-
Convergent evolution of monocyte differentiation in adult skin instructs Langerhans cell identity.Sci Immunol. 2024 Sep 6;9(99):eadp0344. doi: 10.1126/sciimmunol.adp0344. Epub 2024 Sep 6. Sci Immunol. 2024. PMID: 39241057 Free PMC article.
-
Two distinct types of Langerhans cells populate the skin during steady state and inflammation.Immunity. 2012 Nov 16;37(5):905-16. doi: 10.1016/j.immuni.2012.07.019. Immunity. 2012. PMID: 23159228
-
Uncovering the Mysteries of Langerhans Cells, Inflammatory Dendritic Epidermal Cells, and Monocyte-Derived Langerhans Cell-Like Cells in the Epidermis.Front Immunol. 2018 Jul 30;9:1768. doi: 10.3389/fimmu.2018.01768. eCollection 2018. Front Immunol. 2018. PMID: 30105033 Free PMC article. Review.
-
Human epidermal Langerhans cells replenish skin xenografts and are depleted by alloreactive T cells in vivo.J Immunol. 2011 Aug 1;187(3):1142-9. doi: 10.4049/jimmunol.1001491. Epub 2011 Jun 22. J Immunol. 2011. PMID: 21697461
-
Development and homeostasis of 'resident' myeloid cells: the case of the Langerhans cell.Trends Immunol. 2010 Dec;31(12):438-45. doi: 10.1016/j.it.2010.09.003. Epub 2010 Oct 27. Trends Immunol. 2010. PMID: 21030305 Review.
Cited by
-
Hematopoietic Growth Factors Regulate the Entry of Monocytes into the Adult Brain via Chemokine Receptor CCR5.Int J Mol Sci. 2024 Aug 15;25(16):8898. doi: 10.3390/ijms25168898. Int J Mol Sci. 2024. PMID: 39201584 Free PMC article.
-
Microenvironmental and cell intrinsic factors governing human cDC2 differentiation and monocyte reprogramming.Front Immunol. 2023 Jul 19;14:1216352. doi: 10.3389/fimmu.2023.1216352. eCollection 2023. Front Immunol. 2023. PMID: 37539048 Free PMC article.
-
Smad2/4 Signaling Pathway Is Critical for Epidermal Langerhans Cell Repopulation Under Inflammatory Condition but Not Required for Their Homeostasis at Steady State.Front Immunol. 2020 May 7;11:912. doi: 10.3389/fimmu.2020.00912. eCollection 2020. Front Immunol. 2020. PMID: 32457763 Free PMC article.
-
Skin barrier immunity and ageing.Immunology. 2020 Jun;160(2):116-125. doi: 10.1111/imm.13152. Epub 2019 Dec 4. Immunology. 2020. PMID: 31709535 Free PMC article. Review.
-
Tissue-resident, extravascular Ly6c- monocytes are critical for inflammation in the synovium.Cell Rep. 2023 May 30;42(5):112513. doi: 10.1016/j.celrep.2023.112513. Epub 2023 May 18. Cell Rep. 2023. PMID: 37204925 Free PMC article.
References
-
- Capucha T, Mizraji G, Segev H, Blecher-Gonen R, Winter D, Khalaileh A, Tabib Y, Attal T, Nassar M, Zelentsova K, Kisos H, Zenke M, Sere K, Hieronymus T, Burstyn-Cohen T, Amit I, Wilensky A, Hovav AH, Distinct Murine Mucosal Langerhans Cell Subsets Develop from Pre-dendritic Cells and Monocytes. Immunity 43, 369–381 (2015). - PubMed
-
- Schulz C, Gomez Perdiguero E., Chorro L, Szabo-Rogers H, Cagnard N, Kierdorf K, Prinz M, Wu B, Jacobsen SE, Pollard JW, Frampton J, Liu KJ, Geissmann F, A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 336, 86–90 (2012). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
