

Immune function of an angiotensin-converting enzyme against Rice stripe virus infection in a vector insect
- PMID: 31247402
- PMCID: PMC7127076
- DOI: 10.1016/j.virol.2019.05.007
Immune function of an angiotensin-converting enzyme against Rice stripe virus infection in a vector insect
Abstract
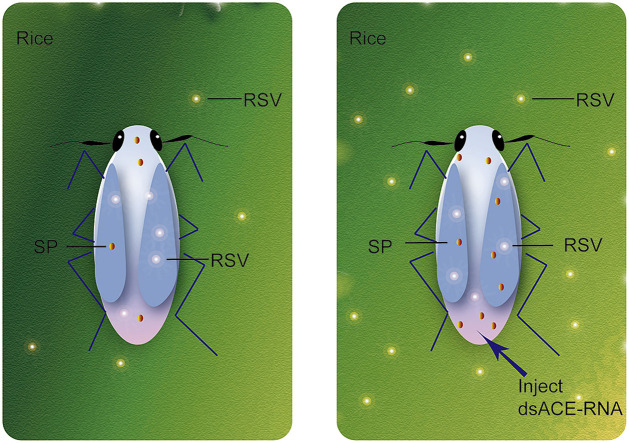
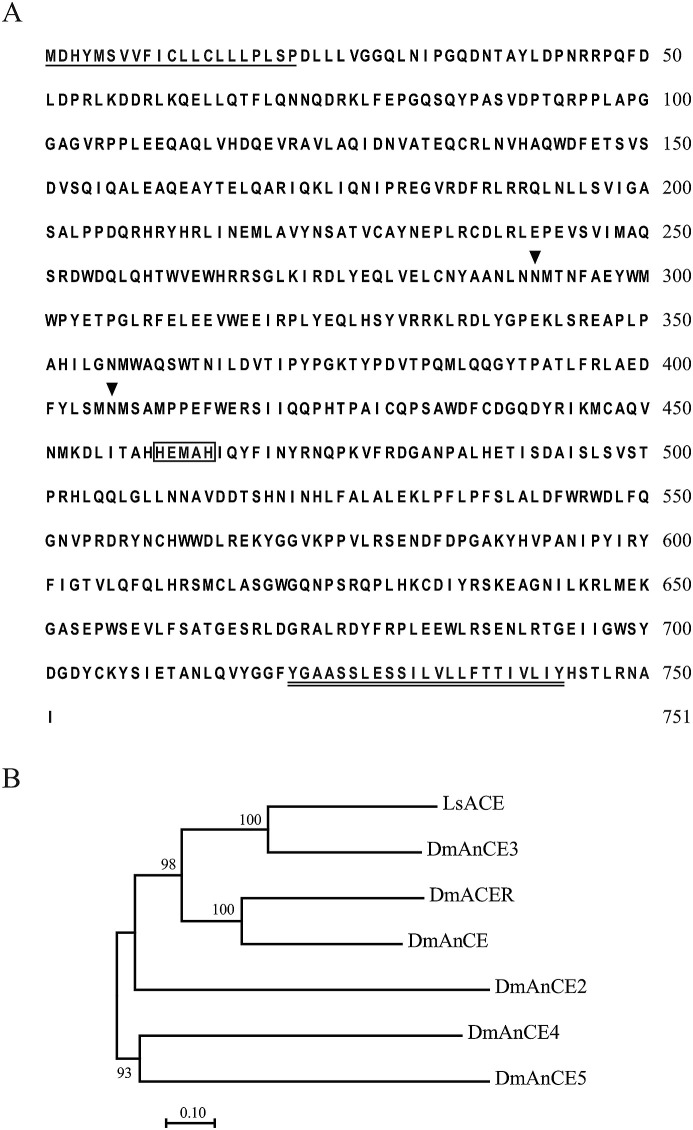
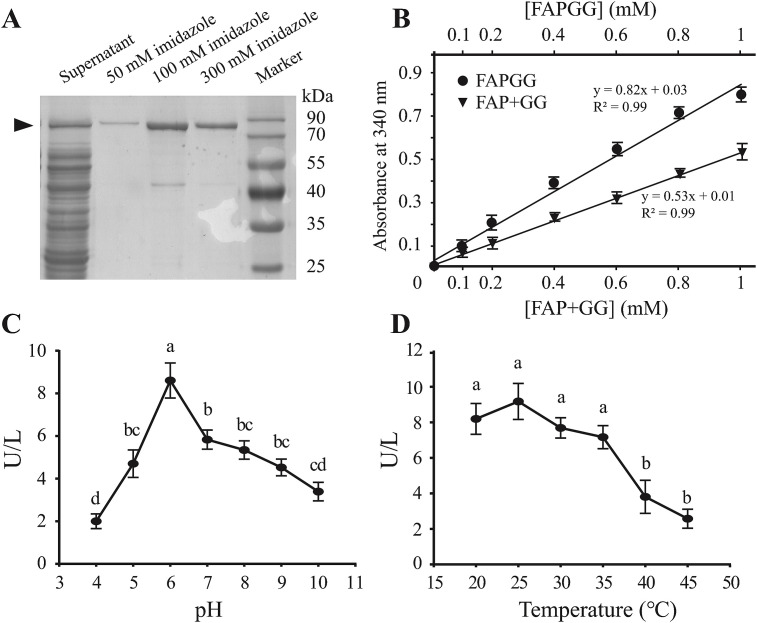
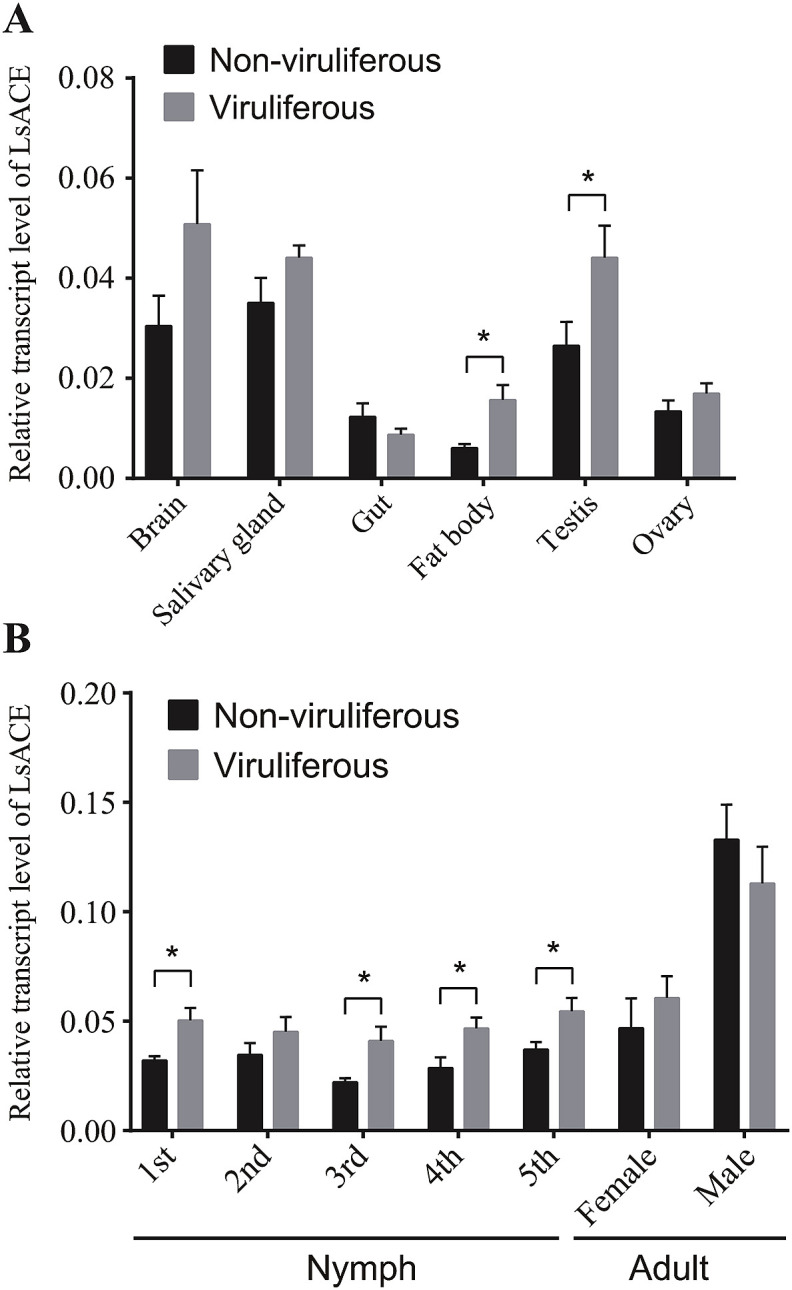
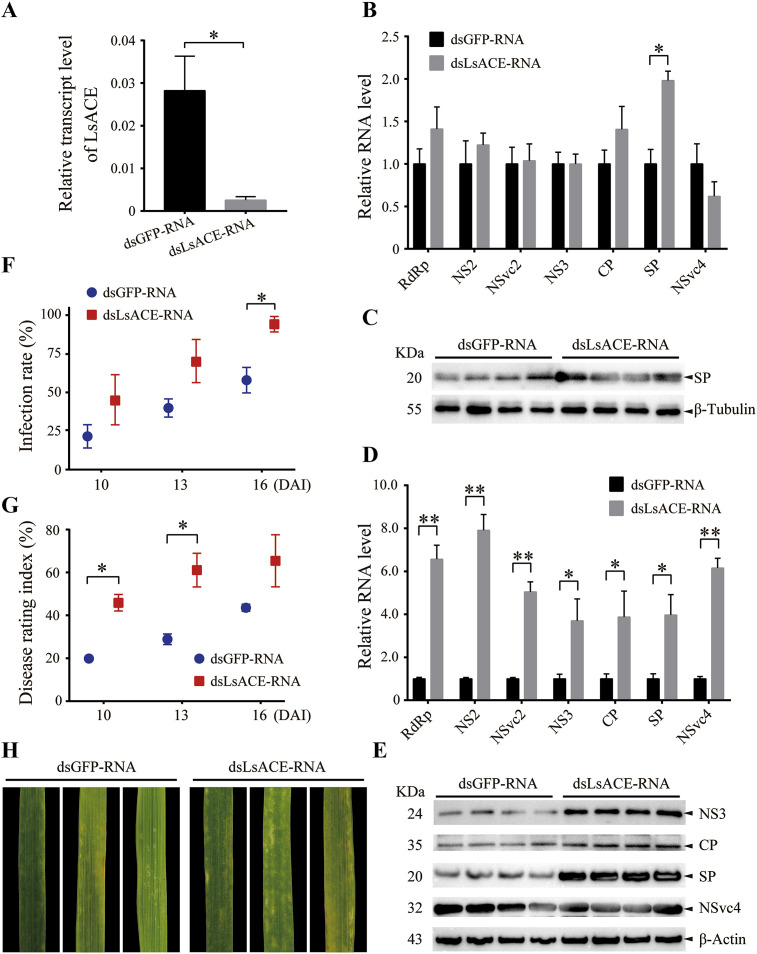
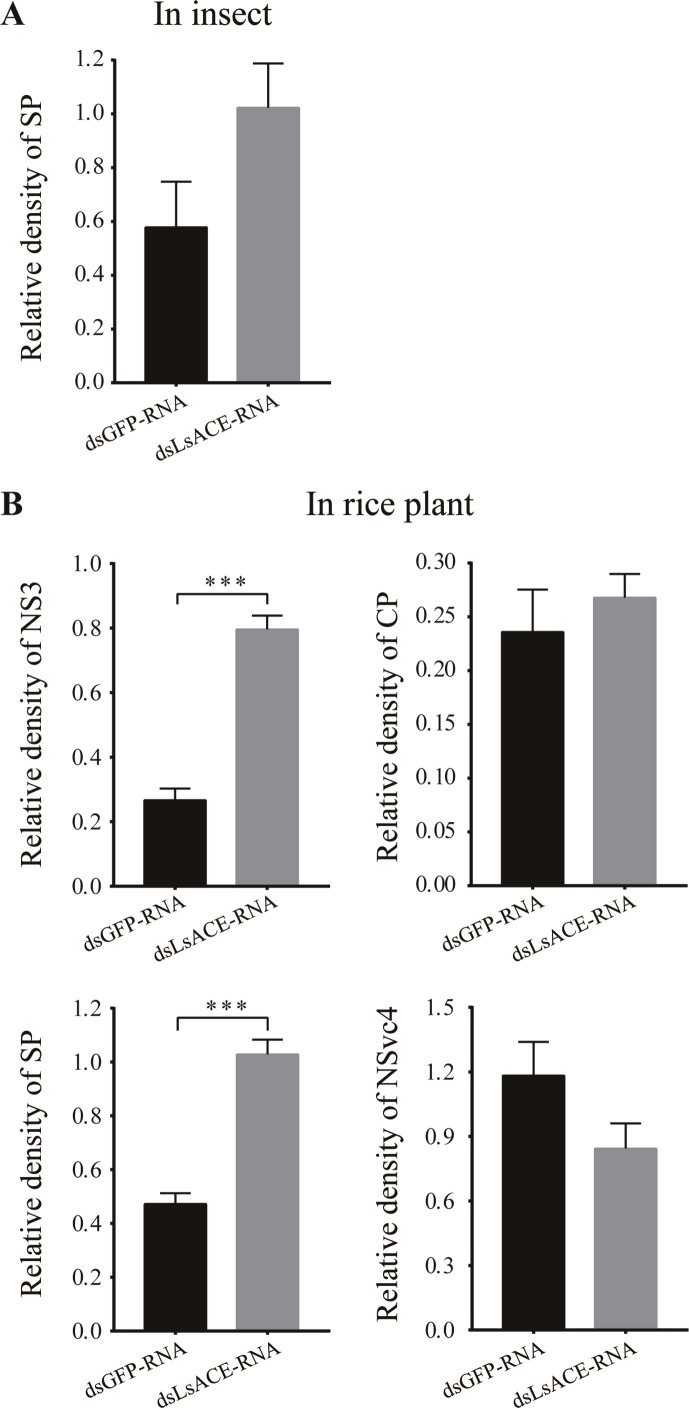
Angiotensin-converting enzyme (ACE) plays diverse roles in the animal kingdom. However, whether ACE plays an immune function against viral infection in vector insects is unclear. In this study, an ACE gene (LsACE) from the small brown planthopper was found to respond to Rice stripe virus (RSV) infection. The enzymatic activities of LsACE were characterized at different pH and temperature. Twenty planthopper proteins were found to interact with LsACE. RSV infection significantly upregulated LsACE expression in the testicle and fat body. When the expression of LsACE in viruliferous planthoppers was inhibited, the RNA level of the RSV SP gene was upregulated 2-fold in planthoppers, and all RSV genes showed higher RNA levels in the rice plants consumed by these planthoppers, leading to a higher viral infection rate and disease rating index. These results indicate that LsACE plays a role in the immune response against RSV transmission by planthoppers.
Keywords: Angiotensin-converting enzyme; Enzyme activity; Immune function; Rice stripe virus; Small brown planthopper.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures

Similar articles
-
Immune responses induced by different genotypes of the disease-specific protein of Rice stripe virus in the vector insect.Virology. 2019 Jan 15;527:122-131. doi: 10.1016/j.virol.2018.11.011. Epub 2018 Nov 27. Virology. 2019. PMID: 30500711
-
Alternative Splicing Landscape of Small Brown Planthopper and Different Response of JNK2 Isoforms to Rice Stripe Virus Infection.J Virol. 2022 Jan 26;96(2):e0171521. doi: 10.1128/JVI.01715-21. Epub 2021 Nov 10. J Virol. 2022. PMID: 34757837 Free PMC article.
-
Rice stripe virus-derived siRNAs play different regulatory roles in rice and in the insect vector Laodelphax striatellus.BMC Plant Biol. 2018 Oct 4;18(1):219. doi: 10.1186/s12870-018-1438-7. BMC Plant Biol. 2018. PMID: 30286719 Free PMC article.
-
The small brown planthopper (Laodelphax striatellus) as a vector of the rice stripe virus.Arch Insect Biochem Physiol. 2023 Feb;112(2):e21992. doi: 10.1002/arch.21992. Epub 2022 Dec 27. Arch Insect Biochem Physiol. 2023. PMID: 36575628 Review.
-
New insights on the transmission mechanism of tenuiviruses by their vector insects.Curr Opin Virol. 2018 Dec;33:13-17. doi: 10.1016/j.coviro.2018.07.004. Epub 2018 Jul 18. Curr Opin Virol. 2018. PMID: 30029017 Review.
Cited by
-
Control of aging by the renin-angiotensin system: a review of C. elegans, Drosophila, and mammals.Front Pharmacol. 2022 Sep 14;13:938650. doi: 10.3389/fphar.2022.938650. eCollection 2022. Front Pharmacol. 2022. PMID: 36188619 Free PMC article. Review.
-
Tachykinin-related peptides modulate immune-gene expression in the mealworm beetle Tenebrio molitor L.Sci Rep. 2022 Oct 14;12(1):17277. doi: 10.1038/s41598-022-21605-6. Sci Rep. 2022. PMID: 36241888 Free PMC article.
-
The nucleocapsid protein of rice stripe virus in cell nuclei of vector insect regulates viral replication.Protein Cell. 2022 May;13(5):360-378. doi: 10.1007/s13238-021-00822-1. Epub 2021 Mar 6. Protein Cell. 2022. PMID: 33675514 Free PMC article.
-
The Bunyavirales: The Plant-Infecting Counterparts.Viruses. 2021 May 6;13(5):842. doi: 10.3390/v13050842. Viruses. 2021. PMID: 34066457 Free PMC article. Review.
References
-
- Aguilar R., Jedlicka A.E., Mintz M., Mahairaki V., Scott A.L., Dimopoulos G. Global gene expression analysis of Anopheles gambiae responses to microbial challenge. Insect Biochem. Mol. Biol. 2005;35:709–719. - PubMed
-
- Baumann R., Casaulta C., Simon D., Conus S., Yousefi S., Simon H.-U. Macrophage migration inhibitory factor delays apoptosis in neutrophils by inhibiting the mitochondria-dependent death pathway. FASEB J. 2003;17:2221–2230. - PubMed
-
- Cheung H.S., Wang F.L., Ondetti M.A., Sabo E.F., Cushman D.W. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. Importance of the COOH-terminal dipeptide sequence. J. Biol. Chem. 1980;255:401–407. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
