

Genome Organization in and around the Nucleolus
- PMID: 31212844
- PMCID: PMC6628108
- DOI: 10.3390/cells8060579
Genome Organization in and around the Nucleolus
Abstract
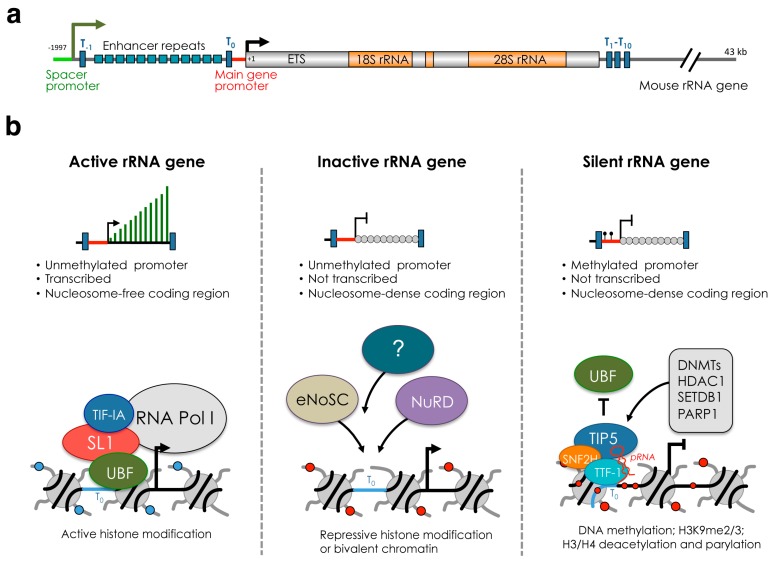
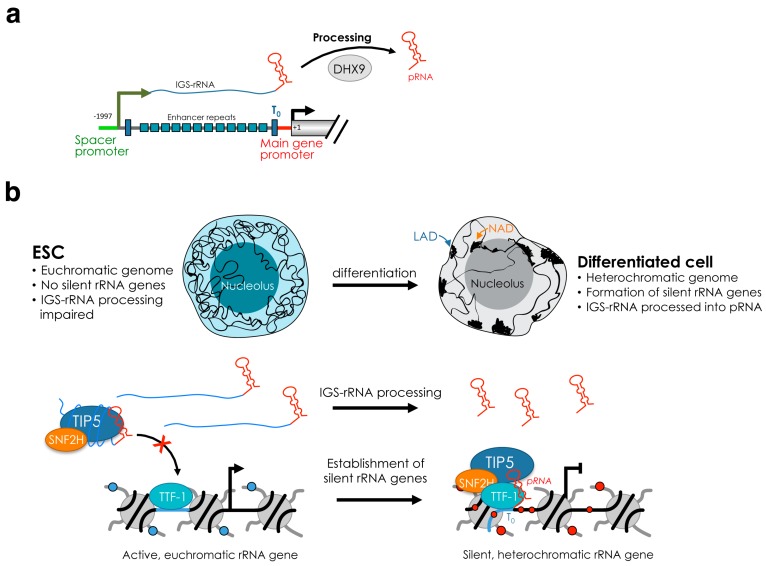
The nucleolus is the largest substructure in the nucleus, where ribosome biogenesis takes place, and forms around the nucleolar organizer regions (NORs) that comprise ribosomal RNA (rRNA) genes. Each cell contains hundreds of rRNA genes, which are organized in three distinct chromatin and transcriptional states-silent, inactive and active. Increasing evidence indicates that the role of the nucleolus and rRNA genes goes beyond the control of ribosome biogenesis. Recent results highlighted the nucleolus as a compartment for the location and regulation of repressive genomic domains and, together with the nuclear lamina, represents the hub for the organization of the inactive heterochromatin. In this review, we aim to describe the crosstalk between the nucleolus and the rest of the genome and how distinct rRNA gene chromatin states affect nucleolus structure and are implicated in genome stability, genome architecture, and cell fate decision.
Keywords: NoRC; Nucleolus; cancer; chromatin; embryonic stem cells; epigenetics; genome architecture; genome instability; lamina-associated domains(LADs); lncRNA; nucleolus-associated domains (NADs); rRNA genes.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Nucleolus and rRNA Gene Chromatin in Early Embryo Development.Trends Genet. 2019 Nov;35(11):868-879. doi: 10.1016/j.tig.2019.06.005. Epub 2019 Jul 18. Trends Genet. 2019. PMID: 31327501 Free PMC article. Review.
-
Regulation and Roles of the Nucleolus in Embryonic Stem Cells: From Ribosome Biogenesis to Genome Organization.Stem Cell Reports. 2020 Dec 8;15(6):1206-1219. doi: 10.1016/j.stemcr.2020.08.012. Epub 2020 Sep 24. Stem Cell Reports. 2020. PMID: 32976768 Free PMC article. Review.
-
Genome organization in and around the nucleolus.Trends Genet. 2011 Apr;27(4):149-56. doi: 10.1016/j.tig.2011.01.002. Epub 2011 Feb 4. Trends Genet. 2011. PMID: 21295884 Review.
-
Plant nucleolar DNA: Green light shed on the role of Nucleolin in genome organization.Nucleus. 2017 Jan 2;8(1):11-16. doi: 10.1080/19491034.2016.1236167. Epub 2016 Sep 20. Nucleus. 2017. PMID: 27644794 Free PMC article. Review.
-
Ribosomal RNA genes shape chromatin domains associating with the nucleolus.Nucleus. 2019 Dec;10(1):67-72. doi: 10.1080/19491034.2019.1591106. Epub 2019 Mar 25. Nucleus. 2019. PMID: 30870088 Free PMC article. Review.
Cited by
-
Stress-Induced Evolution of the Nucleolus: The Role of Ribosomal Intergenic Spacer (rIGS) Transcripts.Biomolecules. 2024 Oct 20;14(10):1333. doi: 10.3390/biom14101333. Biomolecules. 2024. PMID: 39456266 Free PMC article. Review.
-
Regulation of Nucleolar Activity by MYC.Cells. 2022 Feb 7;11(3):574. doi: 10.3390/cells11030574. Cells. 2022. PMID: 35159381 Free PMC article. Review.
-
Personal Perspectives on Plant Ribosomal RNA Genes Research: From Precursor-rRNA to Molecular Evolution.Front Plant Sci. 2021 Dec 21;12:797348. doi: 10.3389/fpls.2021.797348. eCollection 2021. Front Plant Sci. 2021. PMID: 34992624 Free PMC article. Review.
-
Understanding 3D Genome Organization and Its Effect on Transcriptional Gene Regulation Under Environmental Stress in Plant: A Chromatin Perspective.Front Cell Dev Biol. 2021 Dec 8;9:774719. doi: 10.3389/fcell.2021.774719. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34957106 Free PMC article. Review.
-
Lamina-associated domains: peripheral matters and internal affairs.Genome Biol. 2020 Apr 2;21(1):85. doi: 10.1186/s13059-020-02003-5. Genome Biol. 2020. PMID: 32241294 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
