

Helios Deficiency Predisposes the Differentiation of CD4+Foxp3- T Cells into Peripherally Derived Regulatory T Cells
- PMID: 31167776
- PMCID: PMC6615958
- DOI: 10.4049/jimmunol.1900388
Helios Deficiency Predisposes the Differentiation of CD4+Foxp3- T Cells into Peripherally Derived Regulatory T Cells
Abstract
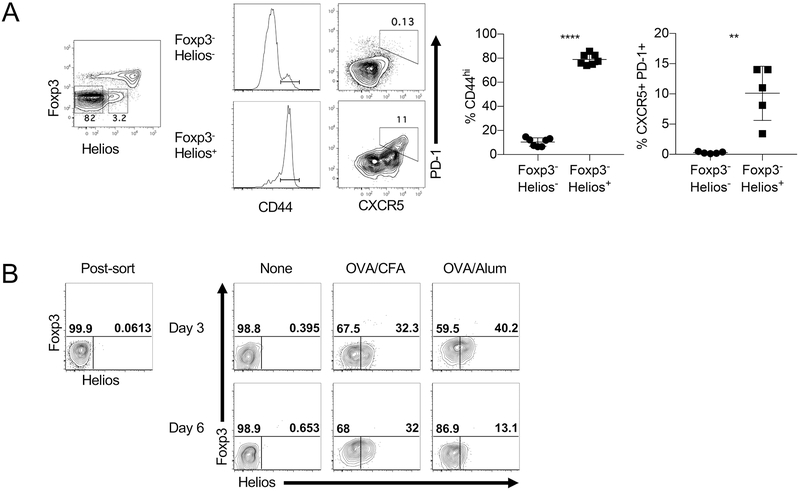
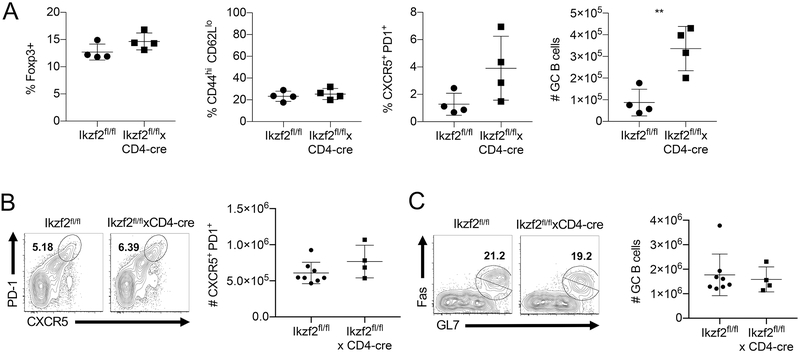
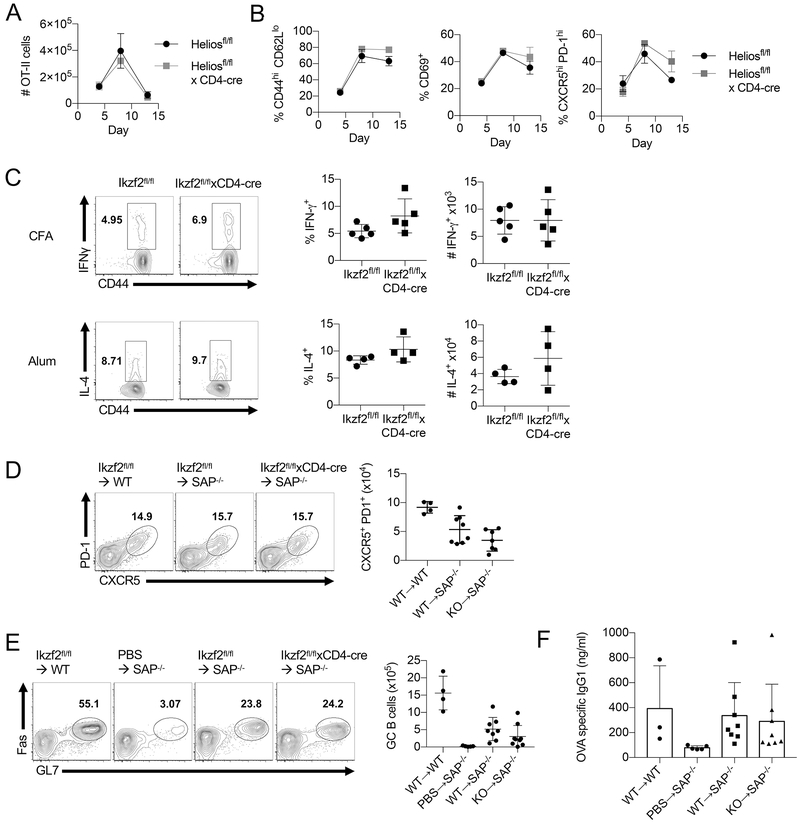
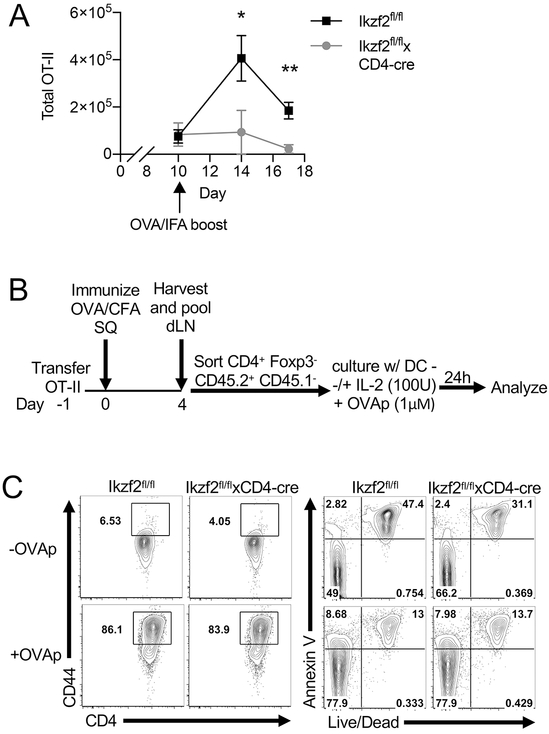
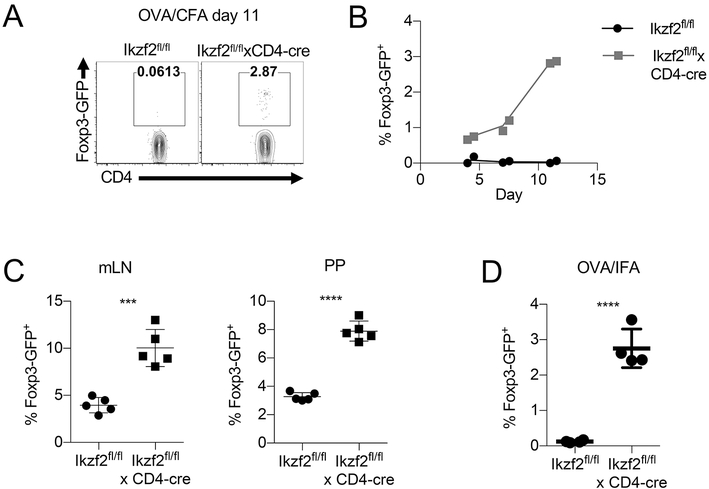
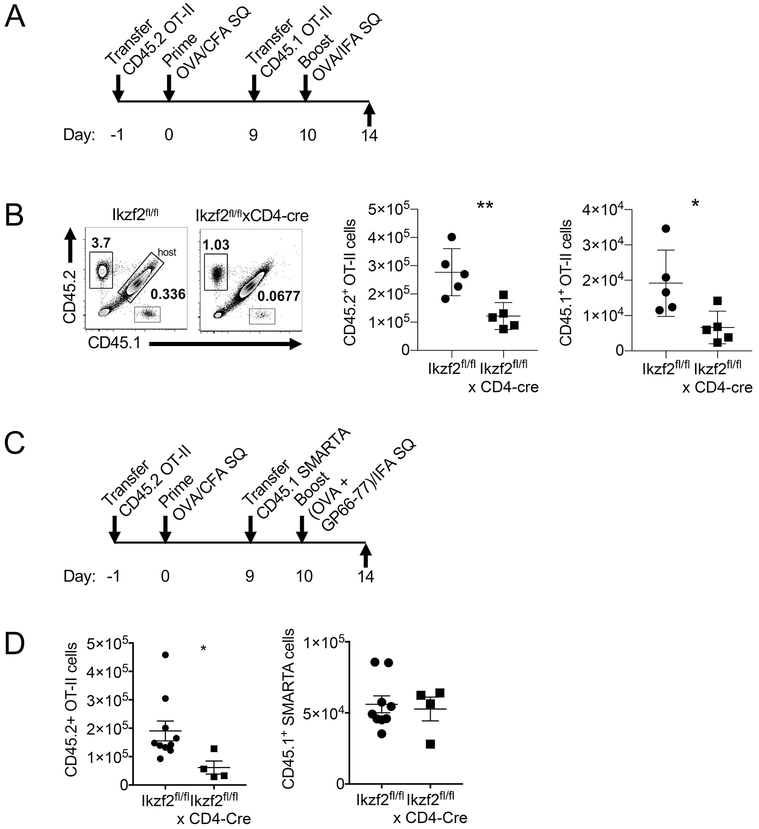
The transcription factor Helios is expressed in a large percentage of Foxp3+ regulatory T (Treg) cells and is required for the maintenance of their suppressive phenotype, as mice with a selective deficiency of Helios in Treg cells spontaneously develop autoimmunity. However, mice with a deficiency of Helios in all T cells do not exhibit autoimmunity, despite the defect in the suppressor function of their Treg cell population, suggesting that Helios also functions in non-Treg cells. Although Helios is expressed in a small subset of CD4+Foxp3- and CD8+ T cells and its expression is upregulated upon T cell activation, its function in non-Treg cells remains unknown. To examine the function of Helios in CD4+Foxp3- T cells, we transferred Helios-sufficient or -deficient naive CD4+Foxp3- TCR transgenic T cells to normal recipients and examined their capacity to respond to their cognate Ag. Surprisingly, Helios-deficient CD4+ T cells expanded and differentiated into Th1 or Th2 cytokine-producing effectors in a manner similar to wild-type TCR transgenic CD4+ T cells. However, the primed Helios-deficient cells failed to expand upon secondary challenge with Ag. The tolerant state of the Helios-deficient memory T cells was not cell-intrinsic but was due to a small population of Helios-deficient naive T cells that had differentiated into Ag-specific peripheral Treg cells that suppressed the recall response in an Ag-specific manner. These findings demonstrate that Helios plays a role in the determination of CD4+ T cell fate.
Figures

Similar articles
-
Helios: still behind the clouds.Immunology. 2019 Nov;158(3):161-170. doi: 10.1111/imm.13115. Epub 2019 Oct 13. Immunology. 2019. PMID: 31517385 Free PMC article. Review.
-
Helios Enhances Treg Cell Function in Cooperation With FoxP3.Arthritis Rheumatol. 2015 Jun;67(6):1491-502. doi: 10.1002/art.39091. Arthritis Rheumatol. 2015. PMID: 25733061
-
Helios+ and Helios- Treg subpopulations are phenotypically and functionally distinct and express dissimilar TCR repertoires.Eur J Immunol. 2019 Mar;49(3):398-412. doi: 10.1002/eji.201847935. Epub 2019 Jan 15. Eur J Immunol. 2019. PMID: 30620397 Free PMC article.
-
Helios Controls a Limited Subset of Regulatory T Cell Functions.J Immunol. 2016 Jan 1;196(1):144-55. doi: 10.4049/jimmunol.1501704. Epub 2015 Nov 18. J Immunol. 2016. PMID: 26582951 Free PMC article.
-
Coexpression of Helios in Foxp3+ Regulatory T Cells and Its Role in Human Disease.Dis Markers. 2021 Jun 22;2021:5574472. doi: 10.1155/2021/5574472. eCollection 2021. Dis Markers. 2021. PMID: 34257746 Free PMC article. Review.
Cited by
-
Nature vs. nurture: FOXP3, genetics, and tissue environment shape Treg function.Front Immunol. 2022 Aug 12;13:911151. doi: 10.3389/fimmu.2022.911151. eCollection 2022. Front Immunol. 2022. PMID: 36032083 Free PMC article. Review.
-
Regulatory T Cells in Severe Persistent Asthma in the Era of Monoclonal Antibodies Target Therapies.Inflammation. 2020 Apr;43(2):393-400. doi: 10.1007/s10753-019-01157-0. Inflammation. 2020. PMID: 31853715 Review.
-
Helios: still behind the clouds.Immunology. 2019 Nov;158(3):161-170. doi: 10.1111/imm.13115. Epub 2019 Oct 13. Immunology. 2019. PMID: 31517385 Free PMC article. Review.
-
Helios Expression in Tumor-Infiltrating Lymphocytes Correlates with Overall Survival of Advanced Gastric Cancer Patients.Life (Basel). 2020 Sep 10;10(9):189. doi: 10.3390/life10090189. Life (Basel). 2020. PMID: 32927747 Free PMC article.
-
Molecular Insights Into Regulatory T-Cell Adaptation to Self, Environment, and Host Tissues: Plasticity or Loss of Function in Autoimmune Disease.Front Immunol. 2020 Sep 15;11:1269. doi: 10.3389/fimmu.2020.01269. eCollection 2020. Front Immunol. 2020. PMID: 33072063 Free PMC article. Review.
References
-
- Fantini MC, Becker C, Monteleone G, Pallone F, Galle PR, and Neurath MF. 2004. Cutting edge: TGF-beta induces a regulatory phenotype in CD4+CD25− T cells through Foxp3 induction and down-regulation of Smad7. J Immunol 172: 5149–5153. - PubMed
-
- Zheng SG, Wang J, Wang P, Gray JD, and Horwitz DA. 2007. IL-2 is essential for TGF-beta to convert naive CD4+CD25− cells to CD25+Foxp3+ regulatory T cells and for expansion of these cells. J Immunol 178: 2018–2027. - PubMed
-
- Cobbold SP, Castejon R, Adams E, Zelenika D, Graca L, Humm S, and Waldmann H. 2004. Induction of foxP3+ regulatory T cells in the periphery of T cell receptor transgenic mice tolerized to transplants. J Immunol 172: 6003–6010. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials