

Evolution of Mechanisms that Control Mating in Drosophila Males
- PMID: 31141679
- PMCID: PMC6646047
- DOI: 10.1016/j.celrep.2019.04.104
Evolution of Mechanisms that Control Mating in Drosophila Males
Abstract
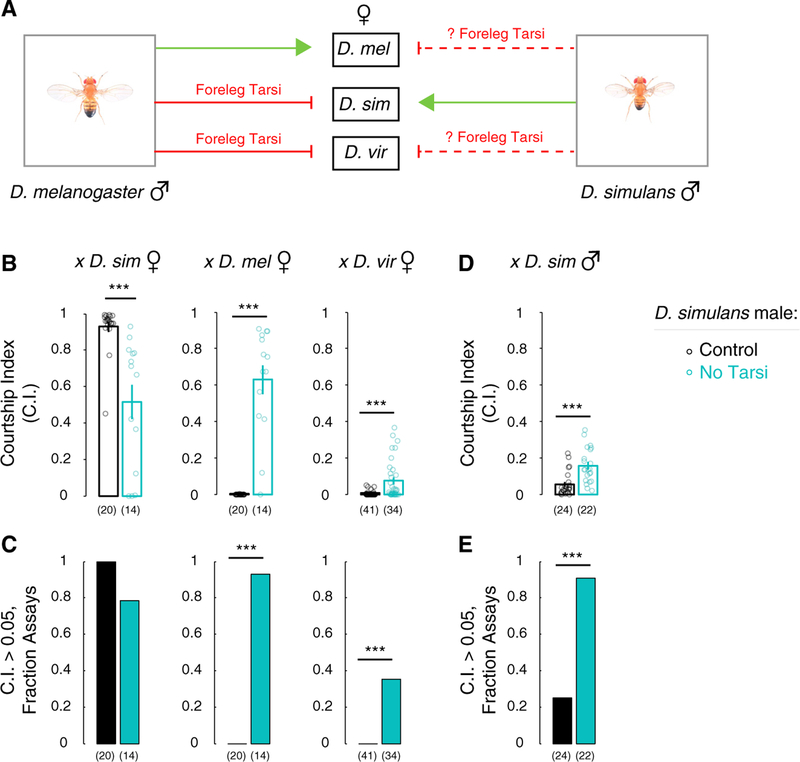
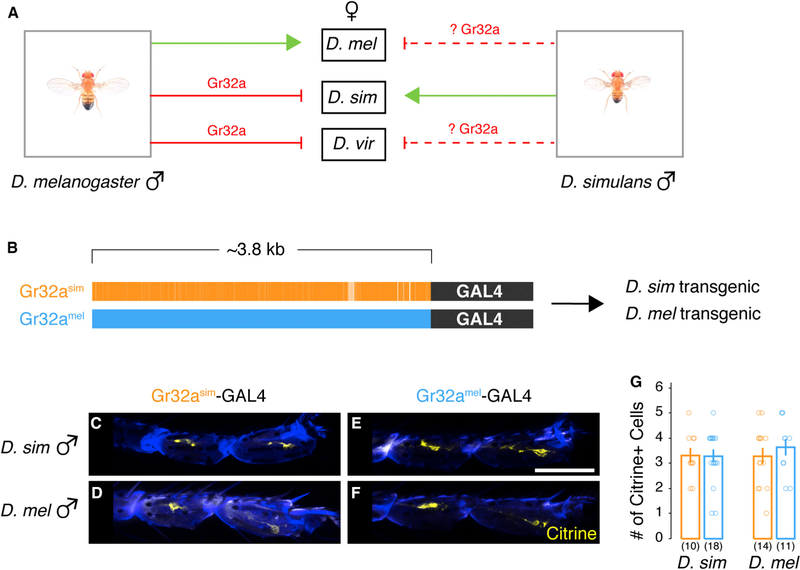
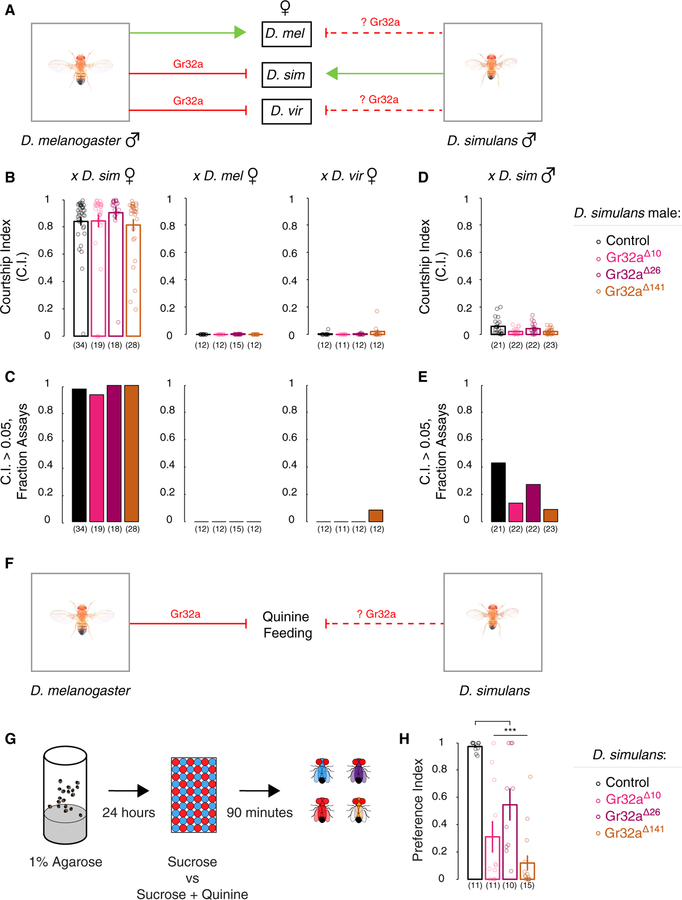
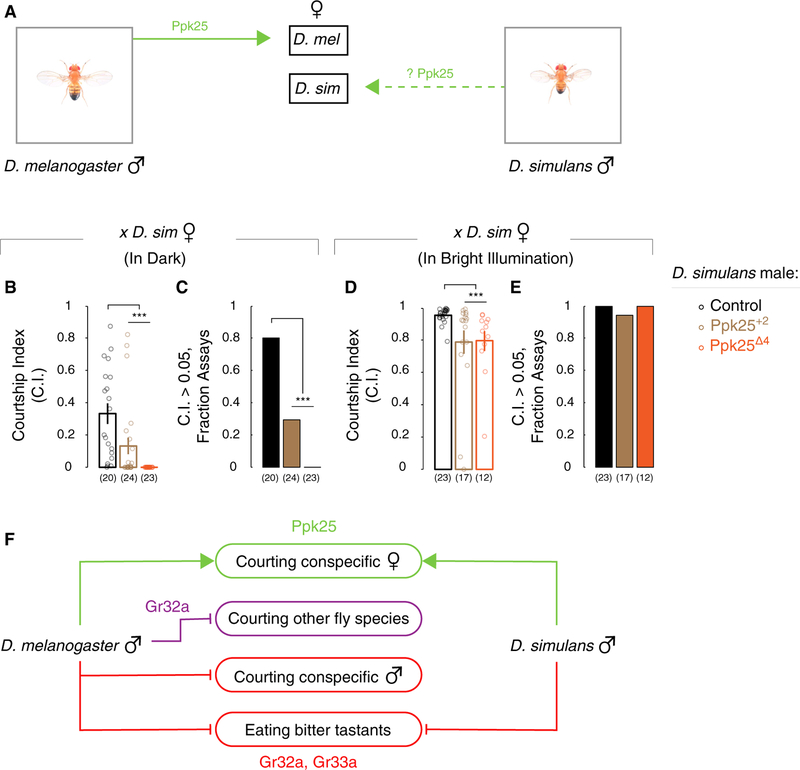
Genetically wired neural mechanisms inhibit mating between species because even naive animals rarely mate with other species. These mechanisms can evolve through changes in expression or function of key genes in sensory pathways or central circuits. Gr32a is a gustatory chemoreceptor that, in D. melanogaster, is essential to inhibit interspecies courtship and sense quinine. Similar to D. melanogaster, we find that D. simulans Gr32a is expressed in foreleg tarsi, sensorimotor appendages that inhibit interspecies courtship, and it is required to sense quinine. Nevertheless, Gr32a is not required to inhibit interspecies mating by D. simulans males. However, and similar to its function in D. melanogaster, Ppk25, a member of the Pickpocket family, promotes conspecific courtship in D. simulans. Together, we have identified distinct evolutionary mechanisms underlying chemosensory control of taste and courtship in closely related Drosophila species.
Keywords: Gr32a; Gr33a; Ppk25; chemosensation; courtship; evolution; mating; pheromones; reproduction; reproductive isolation.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
Genetic and neural mechanisms that inhibit Drosophila from mating with other species.Cell. 2013 Jul 3;154(1):89-102. doi: 10.1016/j.cell.2013.06.008. Epub 2013 Jun 27. Cell. 2013. PMID: 23810192 Free PMC article.
-
Evolution of a central neural circuit underlies Drosophila mate preferences.Nature. 2018 Jul;559(7715):564-569. doi: 10.1038/s41586-018-0322-9. Epub 2018 Jul 11. Nature. 2018. PMID: 29995860 Free PMC article.
-
A Drosophila gustatory receptor essential for aversive taste and inhibiting male-to-male courtship.Curr Biol. 2009 Oct 13;19(19):1623-7. doi: 10.1016/j.cub.2009.07.061. Epub 2009 Sep 17. Curr Biol. 2009. PMID: 19765987 Free PMC article.
-
A taste of the Drosophila gustatory receptors.Curr Opin Neurobiol. 2009 Aug;19(4):345-53. doi: 10.1016/j.conb.2009.07.001. Epub 2009 Aug 5. Curr Opin Neurobiol. 2009. PMID: 19660932 Free PMC article. Review.
-
Evolution of Reproductive Behavior.Genetics. 2020 Jan;214(1):49-73. doi: 10.1534/genetics.119.302263. Genetics. 2020. PMID: 31907301 Free PMC article. Review.
Cited by
-
Adaptive introgression of a visual preference gene.Science. 2024 Mar 22;383(6689):1368-1373. doi: 10.1126/science.adj9201. Epub 2024 Mar 21. Science. 2024. PMID: 38513020 Free PMC article.
-
A modular circuit architecture coordinates the diversification of courtship strategies in Drosophila.bioRxiv [Preprint]. 2023 Sep 17:2023.09.16.558080. doi: 10.1101/2023.09.16.558080. bioRxiv. 2023. Update in: Nature. 2024 Nov;635(8037):142-150. doi: 10.1038/s41586-024-08028-1. PMID: 37745588 Free PMC article. Updated. Preprint.
-
Evolution of central neural circuits: state of the art and perspectives.Nat Rev Neurosci. 2022 Dec;23(12):725-743. doi: 10.1038/s41583-022-00644-y. Epub 2022 Oct 26. Nat Rev Neurosci. 2022. PMID: 36289403 Review.
-
Behavioral Evolution of Drosophila: Unraveling the Circuit Basis.Genes (Basel). 2020 Feb 1;11(2):157. doi: 10.3390/genes11020157. Genes (Basel). 2020. PMID: 32024133 Free PMC article. Review.
-
A modular circuit coordinates the diversification of courtship strategies.Nature. 2024 Nov;635(8037):142-150. doi: 10.1038/s41586-024-08028-1. Epub 2024 Oct 9. Nature. 2024. PMID: 39385031 Free PMC article.
References
-
- Allaire JJ, Cheng J, Xie Y, McPherson J, Chang W, Allen J, Wickham H, Atkins A, Hyndman R, and Arslan R (2017). rmarkdown: Dynamic Documents for R https://cran.r-project.org/web/packages/rmarkdown/index.html.
-
- Bastock M, and Manning A (1955). The courtship of Drosophila melanogaster. Behaviour 8, 85–111.
-
- Billeter J-C, Atallah J, Krupp JJ, Millar JG, and Levine JD (2009). Specialized cells tag sexual and species identity in Drosophila melanogaster. Nature 461, 987–991. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
