

A valve-like mechanism controls desensitization of functional mammalian isoforms of acid-sensing ion channels
- PMID: 31045491
- PMCID: PMC6497441
- DOI: 10.7554/eLife.45851
A valve-like mechanism controls desensitization of functional mammalian isoforms of acid-sensing ion channels
Abstract
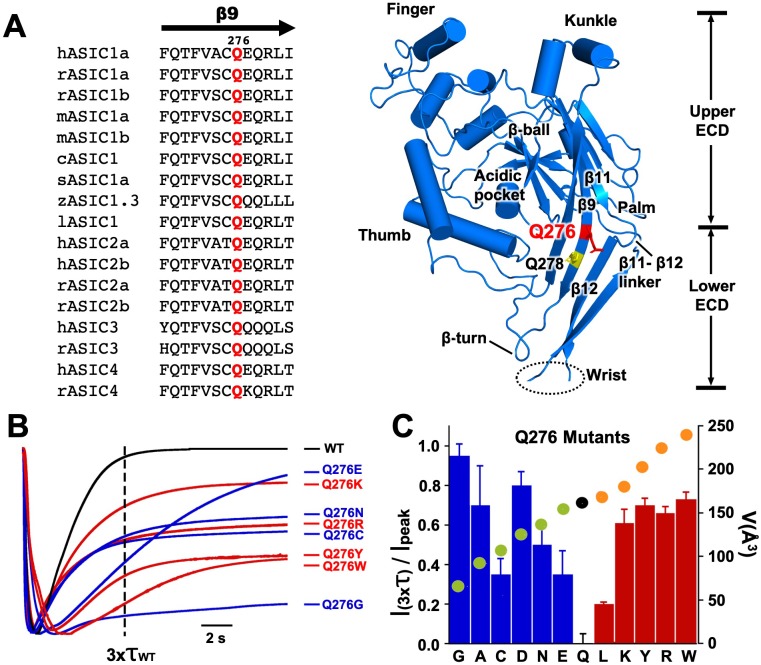
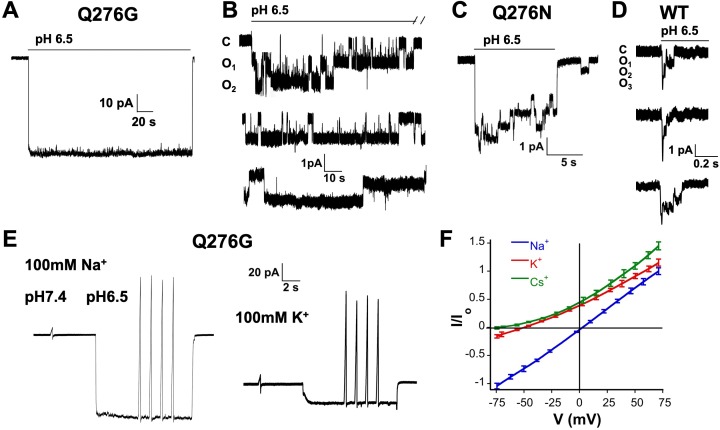
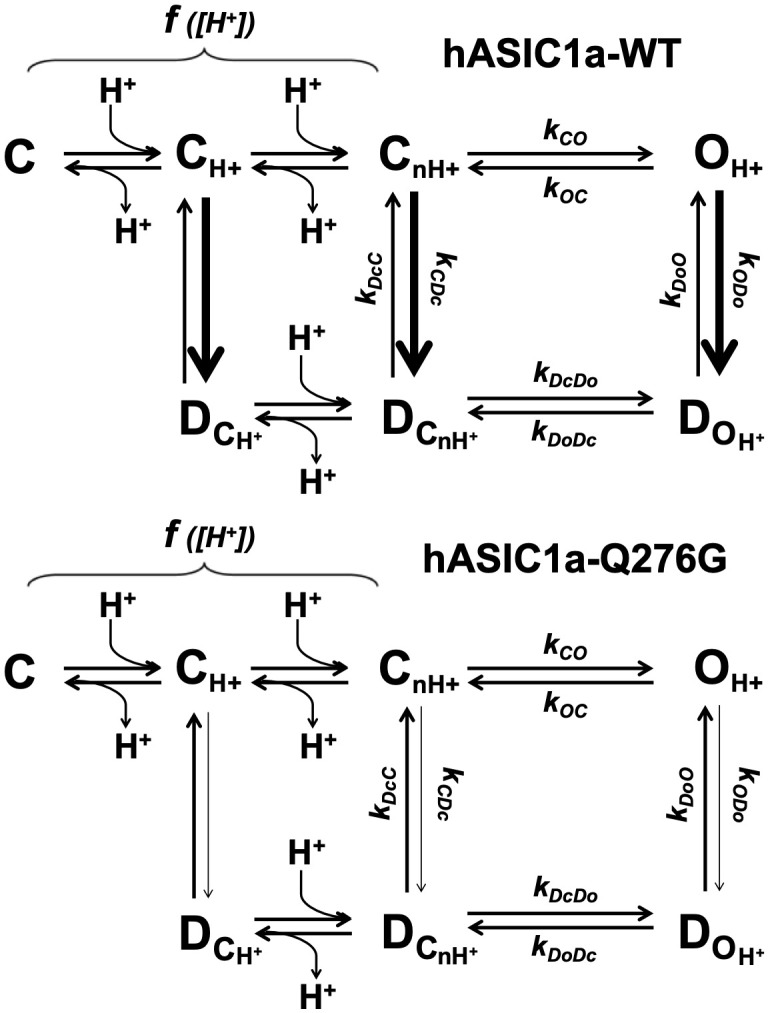
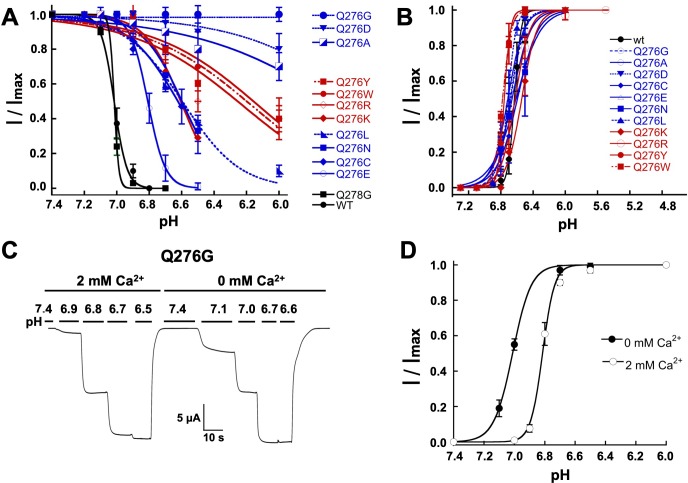
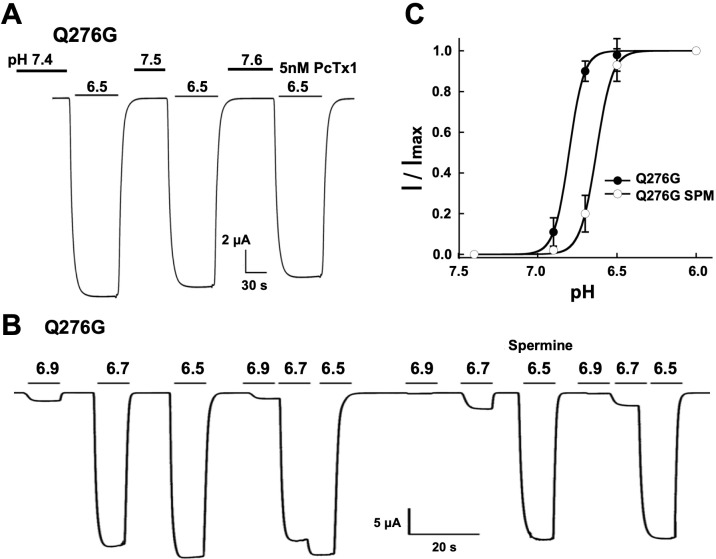
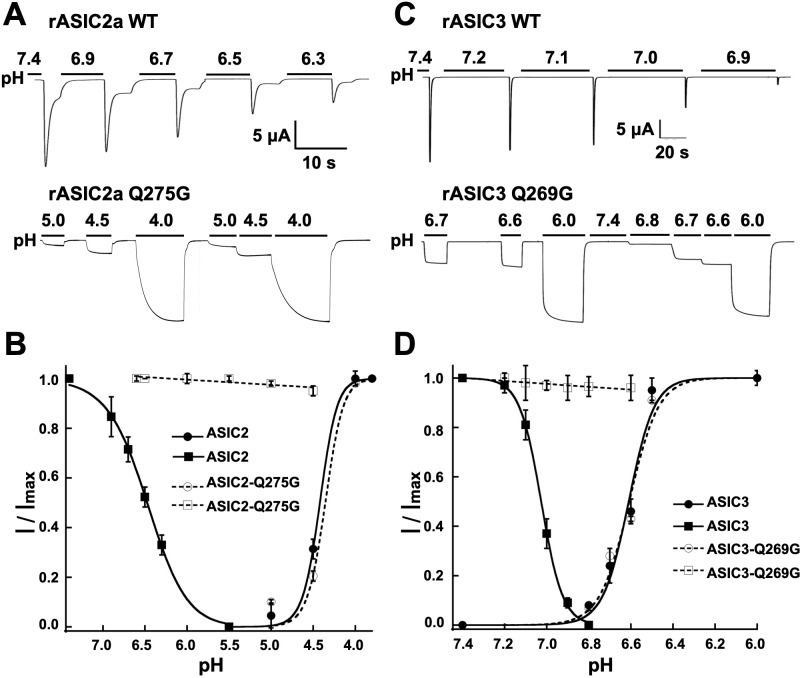
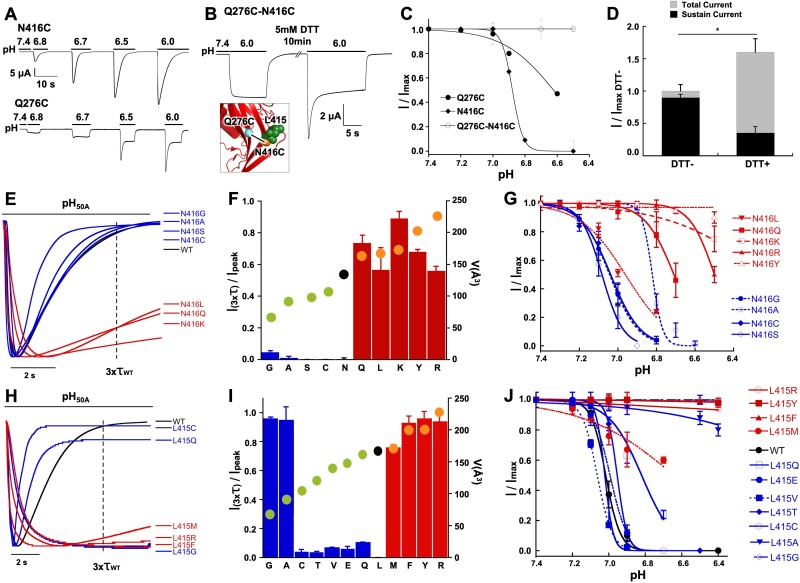
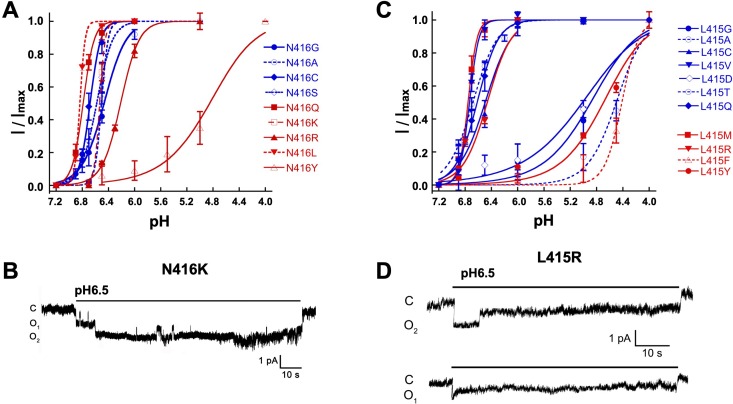
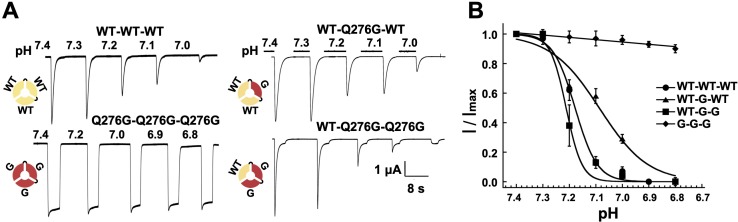
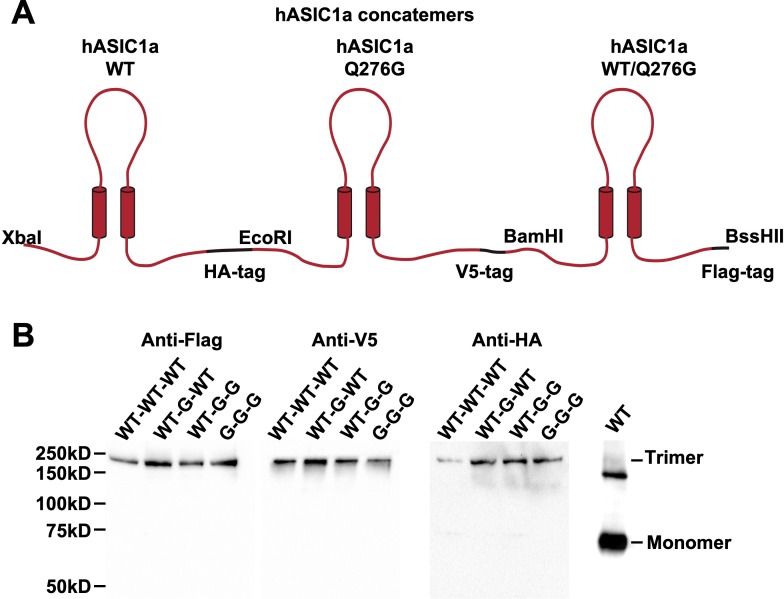
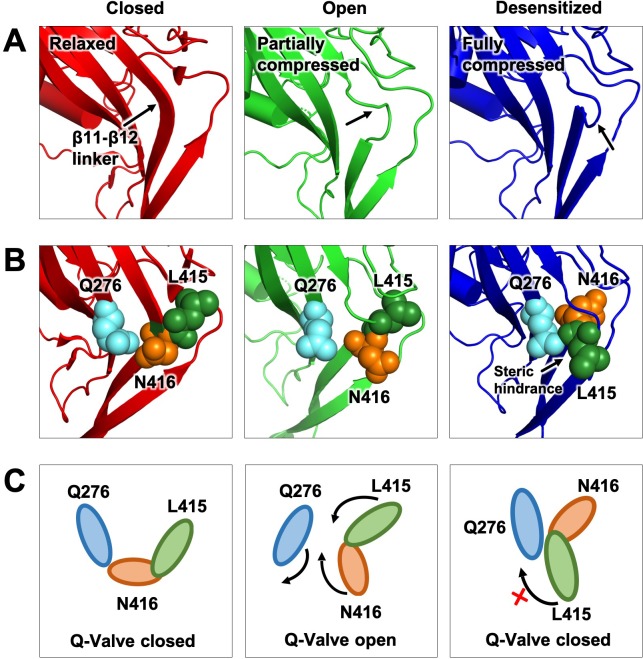
ASICs are proton-gated sodium channels expressed in neurons. Structures of chicken ASIC1 in three conformations have advanced understanding of proton-mediated gating; however, a molecular mechanism describing desensitization from open and pre-open states (steady-state desensitization or SSD) remains elusive. A distinct feature of the desensitized state is an 180o rotation of residues L415 and N416 in the β11- β12 linker that was proposed to mediate desensitization; whether and how it translates into desensitization has not been explored yet. Using electrophysiological measurements of injected Xenopus oocytes, we show that Q276 in β9 strand works with L415 and N416 to mediate both types of desensitization in ASIC1a, ASIC2a and ASIC3. Q276 functions as a valve that enables or restricts rotation of L415 and N416 to keep the linker compressed, its relaxation lengthens openings and leads to sustained currents. At low proton concentrations, the proposed mechanism working in only one of three subunits of the channel is sufficient to induce SSD.
Keywords: ASIC; Q-valve mechanism; b11-b12 linker; desensitization; electrophysiology; neuroscience; xenopus.
© 2019, Wu et al.
Conflict of interest statement
YW, ZC, CC No competing interests declared
Figures

Similar articles
-
Proline substitutions in the ASIC1 β11-12 linker slow desensitization.Biophys J. 2024 Oct 15;123(20):3507-3518. doi: 10.1016/j.bpj.2024.08.016. Epub 2024 Sep 3. Biophys J. 2024. PMID: 39182166
-
Acid-sensing ion channels (ASICs) in mouse skeletal muscle afferents are heteromers composed of ASIC1a, ASIC2, and ASIC3 subunits.FASEB J. 2013 Feb;27(2):793-802. doi: 10.1096/fj.12-220400. Epub 2012 Oct 29. FASEB J. 2013. PMID: 23109675 Free PMC article.
-
The Thumb Domain Mediates Acid-sensing Ion Channel Desensitization.J Biol Chem. 2016 May 20;291(21):11407-19. doi: 10.1074/jbc.M115.702316. Epub 2016 Mar 25. J Biol Chem. 2016. PMID: 27015804 Free PMC article.
-
ASIC and ENaC type sodium channels: conformational states and the structures of the ion selectivity filters.FEBS J. 2017 Feb;284(4):525-545. doi: 10.1111/febs.13840. Epub 2016 Sep 15. FEBS J. 2017. PMID: 27580245 Review.
-
Biophysical properties of acid-sensing ion channels (ASICs).Neuropharmacology. 2015 Jul;94:9-18. doi: 10.1016/j.neuropharm.2014.12.016. Epub 2015 Jan 10. Neuropharmacology. 2015. PMID: 25585135 Review.
Cited by
-
Revealing molecular determinants governing mambalgin-3 pharmacology at acid-sensing ion channel 1 variants.Cell Mol Life Sci. 2024 Jun 17;81(1):266. doi: 10.1007/s00018-024-05276-2. Cell Mol Life Sci. 2024. PMID: 38880807 Free PMC article.
-
Molecular mechanism underlying desensitization of the proton-activated chloride channel PAC.Elife. 2022 Dec 22;11:e82955. doi: 10.7554/eLife.82955. Elife. 2022. PMID: 36547405 Free PMC article.
-
Coupling structure with function in acid-sensing ion channels: challenges in pursuit of proton sensors.J Physiol. 2021 Jan;599(2):417-430. doi: 10.1113/JP278707. Epub 2020 May 20. J Physiol. 2021. PMID: 32306405 Free PMC article. Review.
-
Acid-sensing ion channel 1a modulation of apoptosis in acidosis-related diseases: implications for therapeutic intervention.Cell Death Discov. 2023 Sep 4;9(1):330. doi: 10.1038/s41420-023-01624-6. Cell Death Discov. 2023. PMID: 37666823 Free PMC article. Review.
-
Conformational decoupling in acid-sensing ion channels uncovers mechanism and stoichiometry of PcTx1-mediated inhibition.Elife. 2022 Feb 14;11:e73384. doi: 10.7554/eLife.73384. Elife. 2022. PMID: 35156612 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
